Jan C. Habel und Thomas Schmitt
Zusammenfassung
Die Erhaltung genetischer Diversität ist ein zentrales Ziel eines zeitgemäßen Artenschutzes. Die Ökologie einer Art beeinflusst
deren Populationsstruktur und diese wiederum die genetische Diversität und Differenzierung. So weisen ökologisch spezialisierte Arten,
die meist in kleinen, geographisch isolierten Populationen auftreten, eine vergleichsweise geringe genetische Diversität auf. Dem
gegenüber stehen Arten mit einer vergleichsweise breiten ökologischen Valenz, die in landschaftsübergreifenden Populationsnetzwerken
existieren; durch einen permanenten Austausch von Individuen besitzen diese häufig eine relativ hohe genetische Diversität, die sie
auch weitgehend erhalten können. Aus diesen beiden kontrastierenden Situationen lassen sich zwei unterschiedliche Managementstrategien
für den Naturschutz ableiten: Für ökologisch spezialisierte Arten ist die Erhaltung einer möglichst hohen Habitatqualität essenziell.
Solche Arten können mit ihrer einfachen genetischen Struktur auch langfristig in kleinen isolierten Vorkommen existieren – solange sich
nicht ihre Lebensgrundlage ändert. Dem gegenüber stehen Arten mit einer großen ökologischen Valenz und einer hohen genetischen
Diversität. Diese Arten mit ihren komplexen genetischen Strukturen können nur durch die Erhaltung ihrer Populationsnetzwerke und damit
durch eine hohe Landschaftsdurchlässigkeit erhalten werden. Ein besonderes Augenmerk im Artenschutz muss deshalb auch Arten mit
mittlerer ökologischer Spezialisierung und hoher genetischer Diversität zukommen, da diese nicht dauerhaft in kleinen isolierten
Schutzgebieten erhalten werden können, in besonderer Weise auf Landschaftsdurchlässigkeit angewiesen sind und unter der rasant
fortschreitenden Landschaftshomogenisierung leiden.
Anpassungsfähigkeit – genetische Differenzierung – genetischer Flaschenhals – Genfluss – Habitatisolation – Habitatqualität – Inzuchtdepression – VitalitätAbstract
The preservation of genetic diversity is a central goal of contemporary species conservation. The ecology of a species influences
its population structure, and this in turn influences species‘ genetic diversity and differentiation. For example, ecologically
specialised species, that usually occur in small, geographically isolated populations, have comparatively low genetic diversity. In
contrast, species with a rather broad ecological amplitude exist in cross-country population networks; due to a permanent exchange of
individuals, these often possess a relatively high genetic diversity, that they can also largely maintain through permanent exchanges
of individuals. Two different management strategies for nature conservation can be derived from these two contrasting situations: For
ecologically specialised species, maintaining the highest possible habitat quality is essential. Such species, with their simple
genetic structure, can exist over long terms in small and isolated occurrences – as long as their habitat does not change. On the other
hand, there are species with a high ecological valence and a high genetic diversity. These species with their complex genetic
structures can only be preserved by maintaining their population networks and enabling permanent exchanges of individuals and genes,
and thus by ensuring high landscape permeability. Special attention in species conservation must therefore also be paid to species with
medium ecological specialisation and high genetic diversity, as these cannot be preserved in small isolated protected areas, are
particularly dependent on landscape permeability and suffer from the rapidly progressing landscape homogenisation.
Adaptability – Genetic differentiation – Genetic bottleneck – Gene flow – Habitat isolation – Habitat quality – Inbreeding depression – FitnessInhalt
1 Einleitung
Genetische Diversität ist eine
der wesentlichen Grundvoraussetzungen für die Reaktionsfähigkeit von Organismen auf ihre Umwelt. Sie vergrößert die Variabilität innerhalb
einer Art und erhöht hierdurch auch die Anpassungsfähigkeit an Umweltveränderungen, erlaubt aber auch, auf andere Organismen wie
Prädatoren oder Krankheitserreger zu reagieren (Reed, Frankham 2003). Besonders in Zeiten rascher
globaler und lokaler Veränderungen infolge des Klima- und Landnutzungswandels ist deshalb ein hohes Maß genetischer Variabilität
vorteilhaft für die Anpassungsfähigkeit und damit das Überleben von Arten.
Kasten 1: Glossar.
Box 1: Glossary.
Allel: Die unterschiedlichen Varianten eines Gens an einer bestimmten Stelle (Genort) auf einem Chromosom nennt man Allele.
Sie beeinflussen die Ausprägung eines bestimmten Merkmals (z. B. Blütenfarbe).
Barcoding: Wie bei einem Strichcode auf einer Ware im Laden kann durch die Sequenzierung sogenannter Barcoding-Gene (bei
Tieren oft COI) die Art ermittelt werden.
Dispersal: Alle Lebewesen breiten sich zu einem gewissen Grad aus, dieser unterscheidet sich jedoch zwischen Arten und auch
zwischen den Individuen innerhalb einer Art. Diese Ausbreitung bezeichnet man als Dispersal.
eDNA: Der Begriff steht für Umwelt-DNA (engl.: environmental deoxyribonucleic acid). Hierfür werden Umweltproben genommen
(wie etwa Wasser aus einem Bach) und über die hierin enthaltenen DNA-Moleküle wird ermittelt, welche Arten sich in dem entsprechenden
Bach zu diesem Zeitpunkt befunden haben. Diese Methode erleichtert die Untersuchung von Artengemeinschaften sehr.
Endemisch: Auf einen bestimmten Bereich geographisch beschränkt. Eine Art, die nur in einem begrenzten Gebiet vorkommt,
wird als Endemit dieser Region bezeichnet.
Flaschenhalseffekt: Wenn eine Population einen starken Einbruch ihrer Individuenzahlen durchläuft, so hat dies auch häufig
einen deutlichen Einfluss auf den jeweiligen Genpool, der bei einem solchen Ereignis oftmals kleiner wird. Auch die Frequenzen der
unterschiedlichen Allele können sich hierbei teilweise stark verändern. Da der Verlust genetischer Diversität unter den
Bedingungen eines genetischen Flaschenhalses sehr schnell und scheinbar chaotisch erfolgt, können solche Ereignisse die genetische
Anpassung einer Population an die regionalen Bedingungen negativ beeinflussen. Auch schwach negative Gene können hierbei durch
Zufallsprozesse stark an Bedeutung gewinnen, was zu einer Reduktion der Fitness einer Population führen kann.
Genetische Diversität: Individuen können an Genorten gleiche oder unterschiedliche genetische Information (= Allele)
besitzen. Je mehr unterschiedliche genetische Informationen in einer Population oder Art gefunden werden, desto höher ist ihre
genetische Diversität. Diese kann mit unterschiedlichen genetischen Analyseverfahren (u. a. Sequenzierung, Mikrosatelliten, Allozyme,
double digest restriction-site associated DNA – ddRAD) untersucht und mit verschiedenen Parametern (u. a. Allelzahlen, Heterozygotie,
Polymorphie) dargestellt werden.
Genetische Drift: Die genetische Zusammensetzung einer Population kann sich über die Zeit ändern; hierbei können sich die
Frequenzen der unterschiedlichen Allele verschieben oder es können auch bestimmte Allele ganz verloren gehen; im
Extremfall findet eine Fixierung auf ein einziges Allel an einem Genort statt. Diese Änderungen unterliegen bei
selektionsneutralen Genen rein dem Zufall der Paarungen zwischen den Individuen. Folglich ist die genetische Drift umso stärker
ausgeprägt, je weniger Individuen eine Population aufbauen. Nur bei einer unendlich großen Population (die es jedoch nicht geben kann)
findet keinerlei genetische Drift statt.
Genfluss: Wenn zwischen Populationen Individuen ausgetauscht werden, die sich dann in der jeweils anderen Population
reproduzieren, werden deren Gene in diese Population eingebracht. Ein solcher Genfluss wirkt der genetischen Verarmung von
Populationen, die durch Flaschenhalseffekte, aber auch durch genetische Drift entstehen kann, entgegen.
Inzuchtdepression: Inzucht ist die Fortpflanzung unter mehr oder weniger eng miteinander verwandten Individuen. Hierbei
wird die Wahrscheinlichkeit erhöht, dass ein Nachkomme von beiden Elternteilen dasselbe nachteilige Gen vererbt bekommt. Daher sind
Nachkommen aus Inzuchtpaarungen oft weniger fit als nicht inzüchtig gezeugte Individuen. Inzucht kann deshalb zu einem Fitnessproblem
für Populationen werden.
Metabarcoding: Wie beim normalen Barcoding werden beim Metabarcoding bestimmte Genorte sequenziert, um eine
Artbestimmung vorzunehmen. Jedoch werden beim Metabarcoding eine Vielzahl von Individuen zusammen analysiert (z. B. die Ausbeute aus
einer Insektenfalle), sodass aus dem Ergebnis Rückschlüsse auf alle in der Probe vorhandenen Arten gezogen werden können.
Monophag (Substantiv: Monophagie): Die Breite des Spektrums der von Tieren akzeptierten Nahrung ist von Art zu Art
unterschiedlich. Es gibt Arten mit einem sehr engen Spektrum (= monophag) und solche mit einem sehr weiten (= polyphag). Bei
phytophagen (= Pflanzen fressenden) Insekten wie den Raupen von Schmetterlingen werden Arten, die nur Pflanzenarten einer Gattung als
Nahrung nutzen, oft als monophag bezeichnet; werden nur Arten einer Pflanzenfamilie als Nahrung genutzt, wird dies oft als oligophag
angegeben. Arten, die Pflanzen aus unterschiedlichen Familien nutzen, gelten als polyphag.
Panmiktisch (Substantiv: Panmixie): Alle Individuen innerhalb einer Population verpaaren sich rein zufallsmäßig
untereinander mit jeweils gleich großer Wahrscheinlichkeit.
Vitalität (engl.: fitness): Populationen und Individuen besitzen unterschiedliche Zustände, die zum einen durch die
Lebensumstände, aber auch genetisch bedingt sind. Die Wahrscheinlichkeit zu überleben und ihre Erbanlagen in die nächste Generation
weiterzugeben, unterscheidet sich somit. Dies wird oft als Vitalität oder Fitness bezeichnet.
Genetische Diversität entsteht durch Mutationen, durch die sich ständig neue genetische Varianten bilden, die sog. Allele.
Diese sind wiederum die Grundlage für vererbbare Unterschiede in der Physiologie, Morphologie, Phänologie und des Verhaltens von
Organismen. Diese Unterschiede, basierend auf genetischer Diversität, sorgen auch dafür, dass sich Arten einnischen und koexistieren
können. Bereits Darwin vermutete, dass durch unterschiedliche ökologische Einnischung und damit die unterschiedliche Nutzung der
vorhandenen Ressourcen zahlreiche Arten gemeinsam, also syntop, in einem Lebensraum vorkommen können (Sundberg 2010). Ein wichtiger Aspekt in diesem Zusammenhang ist die Rekombination genetischer Informationen und die daraus
resultierende Anpassungsfähigkeit (Sundberg 2010). Solche Anpassungsprozesse können
auch erstaunlich schnell erfolgen. So wurde z. B. bei Darwinfinken beobachtet, dass sich innerhalb weniger Generationen ihre
Schnabelmorphologie veränderte. Dies wurde auf die veränderte Verfügbarkeit bestimmter Nahrungsressourcen (im Zuge von Nahrungskonkurrenz
oder sich ändernder klimatischer Bedingungen) zurückgeführt (Grant 1986; Grant, Grant 2003, 2006).
Während genetische Diversität, die die Grundlage für die Anpassung an eine sich wandelnde Umwelt darstellt, durch Rekombination
ständig neu variiert wird, wird sie durch genetische Drift reduziert, die Zufallsprozessen unterliegt. Hiervon sind kleine und/oder
isolierte Populationen besonders stark betroffen. Dieser Effekt ist außerdem umso deutlicher ausgeprägt je stärker die Populationsgrößen
schwanken, wobei nur die Minimalgrößen ausschlaggebend sind, die bei sog. Populationsflaschenhälsen auftreten (Flaschenhalseffekt).
Hierbei geht genetische Diversität schnell verloren. Populationsflaschenhälse treten auch unter ganz natürlichen Bedingungen auf. In den
meisten Fällen wird der durch ein solches Ereignis verarmte Genpool in natürlichen Populationsstrukturen durch Einwanderung von Individuen
aus benachbarten Populationen aufgefrischt. Dies geschieht jedoch nur, wenn andere Populationen derselben Art in erreichbarer Nähe
existieren und die Landschaftsmatrix die Ausbreitung (engl. dispersal) der jeweiligen Art durch eine ausreichende Durchlässigkeit
ermöglicht.
Um die zahlreichen Zusammenhänge der genetischen Diversität und Struktur mit der Populationsstruktur und Persistenz von Arten besser
zu verstehen (Abb. 1), sind genetische Analysen eine Grundvoraussetzung. Umfangreiche
molekulargenetische Datensätze lassen sich mittlerweile schnell und kostengünstig erstellen, sowohl auf Grundlage frisch gesammelten
Materials wie auch zunehmend aus alten Sammlungsbelegen. Mit molekulargenetischen Methoden ist es möglich, die genetische Struktur von
Populationen einzelner Arten zu untersuchen (klassische Populationsgenetik), genetische Analysen einzelner Individuen zur
Artidentifikation durchzuführen (Barcoding) sowie ganze Artengemeinschaften aus einem Lebensraum zu beschreiben (Metabarcoding,
eDNA = Umwelt-DNA).

Abb. 1: Gegensätzliche Beziehungen zwischen Abundanz, genetischer Differenzierung und genetischer Diversität im Gradienten
generalistischer und spezialisierter Arten (aus Habel, Schmitt 2012, modifiziert).
Fig. 1: Contrasting relationships between abundance, genetic differentiation, and genetic diversity of generalist and
specialised species (from
Habel, Schmitt 2012, modified).
Mit der rasanten Weiterentwicklung in der Populationsgenetik (hin zur Populationsgenomik) und der Etablierung hocheffizienter neuer
Markersysteme und statistischer Methoden (eine Übersicht hierzu in Kidner et al. 2021) wurde die
populationsgenetische Dimension ein integraler Bestandteil des internationalen (https://www.cbd.int/convention/) sowie nationalen Arten- und Naturschutzes. Sie hat inzwischen auch im praktischen
Naturschutzmanagement Einzug gehalten (Holderegger, Segelbacher 2016; Holderegger et al. 2019). Umso wichtiger ist es, molekulargenetische Kenngrößen wie genetische
Diversität und Struktur sowie mögliche Zusammenhänge mit der Gefährdung von Arten und Populationen, mit deren Verhalten und ökologischen
Ansprüchen sowie mit der Populationsökologie differenziert zu betrachten. Hierzu werden wir nachfolgend wichtige Zusammenhänge am Beispiel
von Tagfaltern beleuchten und kritisch diskutieren, wobei wir v. a. auf den Unterschied zwischen generalistischen und spezialisierten
Arten eingehen und auch die Vorteile sowie Probleme der genetischen Diversität beleuchten.
2 Zusammenhang zwischen genetischer Diversität, Vitalität und ökologischer Anpassungsfähigkeit von Arten
Zahlreiche Studien zeigen, dass eine große genetische Diversität auch mit hoher Vitalität (engl. fitness) einhergeht (Reed, Frankham 2003; Chapman et al. 2009). In diesem
Zusammenhang ist wichtig zu erwähnen, dass die Abundanz und die genetische Diversität mit zunehmender Spezialisierung von Arten
tendenziell abnehmen und die genetische Differenzierung zwischen Populationen zunimmt (Habel, Schmitt
2012; Abb. 1). Arten und Populationen mit einer hohen genetischen Diversität sind
folglich meist anpassungsfähiger und können daher besser auf Umweltveränderungen reagieren, was bspw. durch die regional unterschiedlichen
Gefährdungsgrade von Tagfaltern in Europa und die mit hohen Gefährdungsgraden negativ korrelierten Diversitäten der Populationen gezeigt
wurde (Schmitt, Hewitt 2004). Genetisch diverse Arten und Populationen können deshalb anthropogen
veränderte Lebensräume oft besser besiedeln und auf neue ökologische Nischen ausweichen. Hierdurch kann der Verlust ursprünglicher
Habitate bspw. durch menschliche Aktivitäten oder die Invasion gebietsfremder Arten besser kompensiert werden.
Demgegenüber sind genetisch verarmte Arten und Populationen meist weniger anpassungsfähig und leiden teilweise unter reduzierter
Vitalität. Ein extremes Beispiel ist die Wiesenotter (Vipera ursinii rakosiensis) in der ungarischen Puszta, wo diese
endemische Unterart kurz vor dem Erlöschen steht. Die letzten Populationen sind genetisch stark verarmt und weisen in vielen
Fällen Missbildungen auf, was auf Inzuchtdepressionen hinweist (Újvári et al. 2002); es
sei jedoch erwähnt, dass ukrainische Tiere aus individuenreichen Populationen auch keine Diversität in der untersuchten mitochondrialen
DNA aufweisen, sehr wohl jedoch im für die Immunantwort so wichtigen MHC (major histocompatibility complex) (Újvári et al. 2005). Dennoch müssen die Zusammenhänge zwischen genetischer Diversität und Vitalität
differenziert betrachtet werden, denn geringe genetische Diversität muss nicht zwangsläufig mit einer Reduktion von Vitalität einhergehen.
So sind Arten und Populationen mit geringer genetischer Diversität bekannt, die über eine hohe Vitalität verfügen und überlebensfähig
sind, solange es zu keinen grundlegenden Änderungen in ihrer Umwelt kommt (z. B. Terrell et al.
2016).
2.1 Ökologisch spezialisierte Tagfalterarten mit geringer genetischer Diversität
Eine Metaanalyse an Wiesenschmetterlingen im südwestlichen Deutschland, in Luxemburg und Nordost-Frankreich zeigte, dass Arten mit
sehr speziellen ökologischen Ansprüchen meist eine vergleichsweise geringe genetische Diversität besitzen, also eine einfache genetische
Struktur aufweisen (Habel et al. 2013). Solche genetisch „armen“ Arten sind meist spezifisch an
spezielle Umweltbedingungen wie bestimmte Habitatstrukturen und Ressourcen angepasst. Durch ihre geringe genetische Diversität muss jedoch
angenommen werden, dass solche Arten nur eingeschränkt auf Veränderungen in ihrer Umwelt reagieren können. Hochspezialisierte Arten
existieren natürlicherweise nur an wenigen, ausgewählten Stellen in der Landschaft, die die für diese Taxa essenziellen und spezifischen
Habitatbedingungen aufweisen. Von Natur aus bestehen hochspezialisierte Arten deshalb meistens aus geographisch voneinander isolierten
Populationen. Die im Allgemeinen einfache genetische Ausstattung, bestehend aus nur wenigen Allelen pro Genort, ist meist von einer
weiteren Reduktion nicht gefährdet. Solche Arten sind folglich sowohl genetisch wie auch ökologisch an ganz bestimmte Habitatbedingungen
und an ein Überleben in geographischer Isolation angepasst.
Ein extremes Beispiel für ein solches Taxon ist der Moselapollo (Parnassius apollo vinningensis), der als endemische Unterart
des Roten Apollos (P. apollo) in wenigen Populationen an der unteren Mosel verbreitet ist (Reinhardt
et al. 2020). Diese Vorkommen waren durch die starken Pestizideinsätze im Weinbau in den 1960er- und 1970er-Jahren sehr stark
bedroht und standen kurz vor dem Erlöschen (Kinkler et al. 1987). Durch starke Reduktion des
Einsatzes von Insektiziden im Weinbau (u. a. das Verbot des Einsatzes von Hubschraubern zur Ausbringung) und die Wiederherstellung vieler
Lebensräume wurde jedoch eine schnelle Erholung der Populationen erreicht, trotz des starken Populationsflaschenhalses (Schmidt 1997). Genetische Untersuchungen heutiger Populationen und von Museumsmaterial zeigten, dass
dieser Populationsflaschenhals keine Auswirkungen auf die populationsgenetische Struktur hatte, denn alle Populationen zurück bis 1895
wiesen für die untersuchten Marker (hier Mikrosatelliten) keinerlei genetische Variabilität auf (Habel
et al. 2009b). Dies steht in Einklang mit der starken ökologischen Spezialisierung dieses Taxons, lässt aber auch verstehen,
warum sich die Populationen so gut erholen konnten. Wegen des Fehlens genetischer Diversität – wahrscheinlich Ergebnis eines
langen natürlichen Reduktionsprozesses – wurde die genetische Zusammensetzung der Populationen durch ihren Flaschenhals nicht verändert
(Habel et al. 2009b). Aktuell sind die Populationen jedoch wieder stark rückläufig, was
gegenwärtig aber mit den klimatischen Veränderungen zusammenhängen könnte (Müller, Hanisch 2020).
An diese sind die genetisch verarmten, hochspezialisierten Populationen vermutlich nicht ausreichend angepasst. Es ist somit möglich, dass
sich heute – anders als in der Vergangenheit – die mit fehlender genetischer Diversität zusammenhängenden Probleme bei dieser Unterart
zeigen.
Ein weiteres Beispiel einer Art, die wahrscheinlich auch aufgrund geringer genetischer Diversität gut angepasst ist an das Überleben
in isolierten Populationen, stellt der Ehrenpreis-Scheckenfalter (Melitaea aurelia; Abb. 2) dar, der in vergleichsweise kleinen und isolierten Kalkmagerrasen stabile Populationen besitzt. Diese verfügen aber nur
über einen kleinen Genpool, der sich auch zwischen den Lebensräumen nicht stark unterscheidet (Habel et al.
2009a). Differenzierter stellt sich das Bild für den Blauschillernden Feuerfalter (Lycaena helle) dar, der in den
Mittelgebirgen Deutschlands und Frankreichs auf Brachflächen mit reichen Beständen von Schlangenknöterich (Bistorta officinalis)
Reliktvorkommen besitzt. Obwohl eine Analyse für das mitochondriale Gen COI keine genetische Diversität für diese Populationen aufwies,
ergaben sich für die oftmals variableren Mikrosatelliten auffallend hohe genetische Diversitäten. Das ist bei dem vorliegenden
Isolationsgrad der Populationen eher erstaunlich. Neben dem starken Verlust von Habitaten und den Effekten des für die Art sehr
problematischen Klimawandels könnte diese genetische Diversität somit ein zusätzlicher Grund für die deutlichen Rückgänge von L.
helle sein, denn genetische Flaschenhälse könnten die populationsgenetischen Strukturen in den Vorkommen bereits nachhaltig gestört
haben (Habel et al. 2014; siehe Abschnitt 3).

Abb. 2: Der Ehrenpreis-Scheckenfalter (Melitaea aurelia) ist eine spezialisierte Art, die aber gut in isolierten
Populationen überleben kann.
Fig. 2: Nickerl's fritillary (Melitaea aurelia) is a specialised species, but can survive well in isolated
populations.
2.2 Tagfalterarten mit breiter ökologischer Amplitude und hoher genetischer Diversität
Dem gegenüber stehen Arten, die eine sehr breite ökologische Amplitude aufweisen. Diese Arten existieren in meist großen,
landschaftsübergreifenden Populationen, besiedeln eine Vielzahl von Lebensräumen und nutzen sehr diverse Ressourcen. Solche Arten sind
genetisch deutlich variabler und es ist davon auszugehen, dass sie über eine vergleichsweise große Anpassungsfähigkeit verfügen. Die große
genetische Diversität wird durch eine hohe Abundanz und einen regen Austausch von Individuen zwischen lokalen Populationen
aufrechterhalten. Typische Tagfalterarten, auf die diese Eigenschaften zutreffen und für die sehr hohe genetische Diversitäten
nachgewiesen wurden, sind bspw. der Kleine Fuchs (Aglais urticae; Vandewoestijne et al.
1999), der Gemeine Bläuling (Polyommatus icarus; Schmitt et al. 2003) und das
Große Ochsenauge (Maniola jurtina; Schmitt et al. 2005). Solche Arten besitzen weitgehend
panmiktische Strukturen über den Raum mit großem Genfluss, der dem Verlust genetischer Diversität entgegenwirkt. Ähnliche
Strukturen werden jedoch auch für Arten mit distinkten Populationen beobachtet, wenn diese meist sehr individuenreich sind und es außerdem
zu häufigem Austausch von Individuen zwischen den Teilpopulationen kommt, wie dies z. B. für den Silbergrünen Bläuling (Polyommatus
coridon) gezeigt wurde (Schmitt, Seitz 2002; Schmitt et al.
2006).
3 Bedeutung genetischer Diversität im Naturschutz- und Artenschutzmanagement
Aus den dargelegten Zusammenhängen zwischen der ökologischen Nische einer Art, ihrer Populationsstruktur und genetischen Diversität
ergeben sich unterschiedliche Schlussfolgerungen für ein erfolgreiches Naturschutz- und Artenschutzmanagement. Hierbei kommt gerade auch
der genetischen Diversität eine hohe Bedeutung zu (Habel, Schmitt 2012, 2018). So besitzen Arten
mit komplexen und spezifischen Habitatansprüchen in den meisten Fällen eine Populationsstruktur mit diskreten, räumlich voneinander
deutlich getrennten Einzelpopulationen und eine einfache genetische Konstitution mit nur geringer Diversität innerhalb und zwischen den
Populationen; die Gefahr der Inzuchtdepression ist deshalb in solchen Fällen reduziert, wie in Abschnitt 2 am Moselapollo ausgeführt. Für den Schutz solcher Arten steht somit die Erhaltung starker lokaler Populationen
in den zur Verfügung stehenden Lebensräumen durch die Förderung einer hohen Habitatqualität im Vordergrund.
Sehr gute Beispiele für solche Arten sind die Ameisenbläulinge (Gattung Phengaris), die zwar große Verbreitungsgebiete
innerhalb der Paläarktis besiedeln, innerhalb derer jedoch alle Arten der Gattung nur sehr lokal auftreten. Diese benötigen
sehr spezifische Lebensraumstrukturen. So nutzen sie eine sehr eingeschränkte Auswahl an Fraßpflanzen. Die Raupen des Dunklen
Wiesenknopf-Ameisenbläulings (P. nausithous) und des Hellen Wiesenknopf-Ameisenbläulings (P. teleius; Abb. 3) sind strikt monophag am Großen Wiesenknopf (Sanguisorba officinalis) und die
weiteren Phengaris-Arten fressen nur an wenigen Pflanzenarten einer einzigen Gattung bzw. Familie. Auch sind die Raupen für ihre
Entwicklung zusätzlich auf bestimmte Wirtsameisen in ausreichenden Dichten angewiesen, die die Raupen adoptieren und in ihre Nester
eintragen (Reinhardt et al. 2020). Obwohl über das gesamte Verbreitungsgebiet eine größere Anzahl
an Wirtsameisenarten akzeptiert wird, ist die Toleranz, welche Ameisen akzeptiert werden, regional sehr eng begrenzt (Tartally et al. 2019). Die Ameisenbläulingsarten verfügen allesamt über eine vergleichsweise geringe
genetische Diversität (Pecsenye et al. 2007). Solange keine grundlegenden Veränderungen in ihren
Habitaten geschehen, sind Population solcher Arten oft in der Lage, in isolierten und auch vergleichsweise kleinen Habitaten mit relativ
geringen Populationsgrößen langfristig zu überleben (Habel, Schmitt 2012).

Abb. 3: Der Helle Wiesenknopf-Ameisenbläuling (Phengaris teleius) ist eine hochgradig spezialisierte Tagfalterart, deren
Raupen zuerst ausschließlich an den Blütenständen des Großen Wiesenknopfs (Sanguisorba officinalis) fressen und
anschließend von Knotenameisen in deren Nester eingetragen werden, wo sie als Sozialparasiten ihre Präimaginalphase beenden.
Fig. 3: The scarce large blue (Phengaris teleius) is a highly specialised butterfly species whose caterpillars first
feed exclusively on the inflorescences of great burnet (Sanguisorba officinalis) and are then carried by Myrmicinae ants into
their nests, where they complete their preimaginal phase as social parasites.
Im Gegensatz dazu stehen Arten mit vergleichsweise hoher genetischer Diversität und generell breiterer ökologischer Amplitude.
Intuitiv würde man also denken, dass diese Arten deshalb ein allgemein geringeres Risiko des Rückgangs aufweisen als die vorgenannten mit
spezifischen Habitatanforderungen und geringer genetischer Diversität. Allerdings kann die populationsgenetische Zusammensetzung bei
Populationen mit einer hohen genetischen Diversität durch Populationsflaschenhälse schnell gestört werden. Schwach negative
Genausprägungen (engl. weakly deleterious genes) können hierbei an Bedeutung gewinnen, was einen negativen Einfluss auf die Vitalität von
Individuen und ganzen Populationen haben kann. Stärkere Flaschenhälse können somit für solche Arten deutlich problematischer als für
spezialisierte Arten sein und ein permanenter Austausch von Individuen zwischen Populationen ist erforderlich, um die hohe genetische
Diversität langfristig zu erhalten.
Das stellt für echte Generalisten, die weit über die Landschaft verbreitet sind, keinerlei Probleme dar. So ist nicht damit zu
rechnen, dass Arten wie der Zitronenfalter (Gonepteryx rhamni; Abb. 4a) oder das Große
Ochsenauge (Maniola jurtina; Abb. 4b) von diesen Ursachen für eine Gefährdung zukünftig
betroffen sein könnten. Für solche Arten ist ein ausreichender Individuenaustausch auf der Landschaftsebene sichergestellt, sogar wenn –
wie im Fall des Zitronenfalters – nur wenige, dafür aber häufige und weit verbreitete Raupenfraßpflanzen genutzt werden. Anders stellt es
sich jedoch für Arten dar, die in der bäuerlichen Kulturlandschaft bis weit ins 20. Jahrhundert hinein als Trivialarten galten, da ihre
Lebensräume in dieser Landschaft ubiquitär waren, wie etwa blütenreiche Wiesen und Weiden. Die für diese Arten erforderlichen Strukturen
sind jedoch mittlerweile sehr stark zurückgegangen, sodass solche ehemaligen Allerweltsarten der Blumenwiesen ebenfalls selten geworden
sind, obwohl viele dieser Arten moderate Generalisten sind mit nur wenigen spezifischen Ansprüche an ihre Lebensräume, die aber nicht in
der modernen ausgeräumten Agrarlandschaft erfüllt werden. Es sind solche Arten, die durch Isolation und Populationsflaschenhälse aktuell
besonders gefährdet sind, denn hierauf waren sie mit ihrer genetischen Ausstattung nicht evolutiv „vorbereitet“.

Abb. 4: a) Der Zitronenfalter (Gonepteryx rhamni) und b) das Große Ochsenauge (Maniola jurtina) sind weit
verbreitet und häufig in Deutschland anzutreffen.
Fig. 4: a) The brimstone (Gonepteryx rhamni) and b) the meadow brown (Maniola jurtina) are widespread and common
in Germany.
Ein eindrückliches Beispiel für dieses Phänomen stellt die Berghexe (Chazara briseis; Abb. 5) dar. Ihre Raupen besitzen durchaus hohe Ansprüche an das Mikroklima und die Vegetationsstrukturen (Kadlec et al.
2009); deshalb war die Berghexe in Deutschland auch in der Vergangenheit auf spezifische Bereiche oft mit intensiver
Schafbeweidung beschränkt (Reinhardt et al. 2020). Trotzdem war die Art früher im
Bereich der Stadt Prag auf Kalkmagerrasen so häufig, dass sie von der dortigen Bevölkerung übersetzt den Namen Prager Schmetterling bekam.
Mittlerweile ist die Art hier ausgestorben und kommt in Böhmen nur noch ganz im Norden des Gebiets auf Steppenrasen des Böhmischen
Mittelgebirges (tschech. České středohoří) vor (Vrba et al. 2021). Diese Populationen wurden
sowohl durch Fang-Wiederfang als auch populationsgenetisch untersucht. Dies zeigte, dass sich die Individuen stark zwischen den einzelnen
Populationen austauschen und auch eine hohe genetische Diversität besitzen (Kadlec et al. 2010).
Die deutliche Isolation und schleichende Verschlechterung der Habitatqualität haben wohl zu einem unzureichenden Austausch von Individuen
zwischen den Populationen, zur Reduzierung der effektiven Populationsgrößen, hieraus resultierend eventuell zu einer negativen
Beeinflussung der genetischen Komposition und zu Inzuchtdepression geführt. Hierauf ist eventuell auch die deutlich reduzierte Vitalität
der Individuen bei der Zucht zurückzuführen, die tschechische Kollegen festgestellt haben (Martin Konvička, pers. Mitt.). Um die letzten
böhmischen Vorkommen zu retten, sollen zur genetischen „Sanierung“ der Populationen nun Individuen aus Sachsen-Anhalt eingebracht werden,
wo sich die letzten stabilen Populationen Deutschlands befinden (Reinhardt et al. 2020). Auch
wird versucht, die Habitatqualitäten wieder zu verbessern, da wegen der hohen Lebensraumansprüche der Raupen (Kadlec et al. 2009) eine mangelnde Habitatqualität neben genetischen Aspekten sicherlich auch ein
wesentlicher Grund für den Zusammenbruch der Populationen war.

Abb. 5: Die Berghexe (Chazara briseis) ist in den letzten Jahrzehnten in Mitteleuropa sehr stark zurückgegangen. Auch
aufgrund ihrer hohen genetischen Diversität wird vermutet, dass sie empfindlich auf die Isolierung ihrer Lebensräume reagiert.
Fig. 5: The hermit (Chazara briseis) has declined sharply in Central Europe in recent decades. Also due to its high
genetic diversity, it is suspected that it is sensitive to habitat isolation.
Auch wenn es sich bei der Berghexe um einen sehr drastischen Fall handelt, so ist doch davon auszugehen, dass etliche weitere Arten
in Deutschland, die keine sehr hohen Anforderungen an ihre Lebensräume stellen, aber trotzdem zumindest regional stark zurückgehen, aus
ähnlichen Gründen verschwinden. So gehen wir z. B. bei den regionalen oder sogar bundesweiten Rückgängen vieler Scheckenfalter (Gattung
Melitaea), aber auch des Perlgrasfalters (Coenonympha arcania; Abb. 6) davon
aus, dass diese neben den ökologischen auch populationsgenetische Gründe haben (siehe Abschnitt 4). Gerade für solche Arten mit mittlerem Spezialisierungsniveau, aber der Notwenigkeit der Ausbildung großer,
genetisch vernetzter Populationen, um ihre Genpools aufrecht zu erhalten, müssen Naturschutzmaßnahmen im Vordergrund stehen, die die
Durchlässigkeit der Landschaft prioritär berücksichtigen, z. B. durch die Schaffung von Korridoren und Trittsteinbiotopen.

Abb. 6: Der Perlgrasfalter (Coenonympha arcania) ist in vielen Regionen Deutschlands noch relativ häufig, geht aber
vielerorts deutlich zurück, was v. a. durch Habitatfragmentierung begründet sein dürfte.
Fig. 6: The pearly heath (Coenonympha arcania) is still relatively common in many regions of Germany, but is declining
significantly in many places, which may be due primarily to habitat fragmentation.
4 Fazit
Die dargelegten Zusammenhänge zwischen ökologischer Amplitude und Verhalten, Populationsstrukturen und genetischen Eigenschaften
führen zu zwei kontrastierenden naturschutzfachlichen Managementstrategien:
1. Erhaltung einer hohen Lebensraumqualität für spezialisierte und standorttreue Arten mit einer vergleichsweise einfachen
genetischen Struktur und 2. Förderung von Landschaftsdurchlässigkeit und eines Netzwerks von Habitaten, um einen regen Austausch von Individuen zu
ermöglichen und auf diese Weise Arten mit einer zum Teil sehr diversen und komplexen genetischen Diversität zu erhalten.
Hohe genetische Diversität befördert die Fähigkeit zur Anpassung einer Art an sich wandelnde Umweltbedingungen, was v. a. in Zeiten
rasch ablaufender anthropogener Veränderungen wie der landwirtschaftlichen Intensivierung bedeutsam ist. Allerdings kann eine hohe
genetische Diversität mit komplexen Strukturen nur durch konstant hohe Populationsgrößen ohne starke Schwankungen oder durch permanenten
Austausch von Individuen aufrechterhalten werden. Somit ergeben sich große Herausforderungen für das Management von Arten mit einer
komplexen und hohen genetischen Diversität. Dies kann anspruchsvoller sein als die Erhaltung von Arten und Populationen, die in
geographischer Isolation über lange Zeit überleben können und deshalb genetisch an eine solche Situation angepasst sind. Zu diesen
sensiblen Arten gehören in Mitteleuropa viele, die ursprünglich in heterogenen Agrarlandschaften mit mageren Wiesen, Brachen, Hecken und
strukturierten Waldrändern weit verbreitet waren oder zumindest in dichten Populationsnetzwerken existierten, jedoch heute stark unter der
zunehmenden Isolation der verbliebenen Habitate leiden. Es sind genau diese Arten mittlerer ökologischer Spezialisierung, die durch den
Landnutzungswandel der letzten Jahrzehnte besonders stark von genetischer Verarmung und Inzuchtdepressionen betroffen sind (Abb. 7), da in weiten Bereichen keine Auffrischung des Genpools durch einen regen Austausch
von Individuen mehr gewährleistet ist.
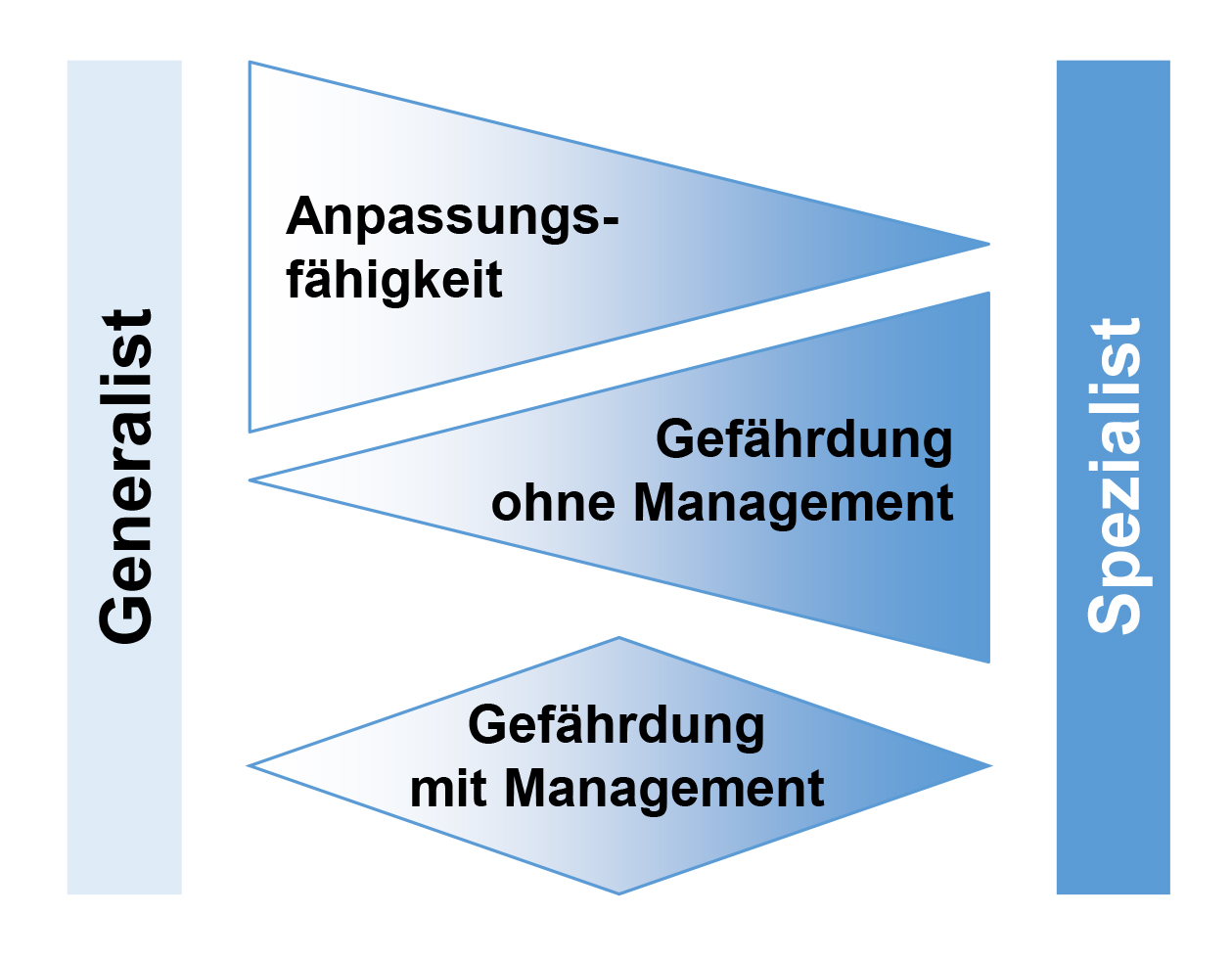
Abb. 7: Gegensätzliche Kohärenz der Anpassungsfähigkeit von Arten und ihrer Gefährdung (ohne Erhaltungsmaßnahmen/Management) im
Gradienten zwischen Generalisten und Spezialisten. Die Anfälligkeit mit Erhaltungsmaßnahmen/Management ist am höchsten für
intermediäre Taxa und am niedrigsten für Generalisten und Spezialisten (aus Habel, Schmitt
2012, modifiziert).
Fig. 7: Opposite coherence of species adaptability and vulnerability (without conservation measures) in the gradient among
generalists and specialists. Vulnerability with conservation measures is highest for intermediate taxa and lowest for generalists
and specialists (from
Habel, Schmitt 2012, modified).
In solchen Fällen ist – wie im Extremfall der Berghexe exemplarisch herausgearbeitet (siehe Abschnitt 3) – die hohe genetische Diversität nicht nur eine positive Voraussetzung für einen anpassungsfähigen Organismus,
sondern kann auch eine Herausforderung darstellen, wenn es um die langfristige Erhaltung solcher Arten mit ihren komplexen genetischen
Strukturen geht (Habel, Schmitt 2012, 2018). Die
dargelegten Beispiele unterstreichen, dass ein differenziertes Verständnis von genetischer Diversität auf der einen Seite und Vitalität
von Organismen und lokalen Populationen auf der anderen Seite essenziell für ein erfolgreiches Management ist und deshalb auch unbedingt
im Naturschutz berücksichtigt werden muss.
5 Literatur
↑
Chapman J.R., Nakagawa S. et al. (2009): A quantitative review of heterozygosity-fitness correlations in animal populations. Molecular Ecology 18(13):
2.746 – 2.765.
↑
Grant P.R. (1986): Ecology and evolution of Darwin's finches. Princeton University Press. Princeton:
492 S.
↑
Grant P.R., Grant B.R. (2006): Evolution of character displacement in Darwin's finches. Science 313(5.784):
224 – 226.
↑
Grant R.B., Grant P.R. (2003): What Darwin's finches can teach us about the evolutionary origin and regulation of
biodiversity. BioScience 53(10): 965 – 975.
↑
Habel J.C., Meyer M., Schmitt T. (2009a): The genetic consequence of differing ecological demands of a generalist and a specialist butterfly species.
Biodiversity and Conservation 18(7): 1.895 – 1.908.
↑
Habel J.C., Meyer M. et al. (2014): The molecular biogeography of the violet copper Lycaena helle.
In: Habel J.C: Jewels in the mist: A synopsis on the endangered violet copper butterfly Lycaena helle. Pensoft. Sofia:
111 – 124.
↑
Habel J.C., Rödder D. et al. (2013): The genetic signature of ecologically different grassland Lepidopterans. Biodiversity and Conservation 22(10):
2.401 – 2.411.
↑
Habel J.C., Schmitt T. (2012): The burden of genetic diversity. Biological Conservation 147(1): 270 – 274.
↑
Habel J.C., Schmitt T. (2018): Vanishing of
the common species: Empty habitats and the role of genetic diversity. Biological Conservation 218: 211 – 216.
↑
Habel J.C., Zachos F.E. et al. (2009b): Unprecedented long-term genetic monomorphism in an endangered relict butterfly species.
Conservation Genetics 10(6): 1.659 – 1.665.
↑
Holderegger R., Balkenhol N. et al. (2019): Conservation genetics: Linking science with practice. Molecular Ecology 28(17): 3.848 – 3.856.
↑
Holderegger R., Segelbacher G. (Hrsg.) (2016): Naturschutzgenetik. Ein Handbuch für die Praxis. Haupt.
Bern: 247 S.
↑
Kadlec T., Vrba P., Konvicka M. (2009): Microhabitat requirements of caterpillars of the critically endangered
butterfly Chazara briseis (L.) (Nymphalidae: Satyrinae) in the Czech Republic. Nota
Lepidopterologica 32(1): 39 – 46.
↑
Kadlec T., Vrba P. et al. (2010): Tracking the decline of the once-common butterfly: Delayed oviposition, demography
and population genetics in the hermit Chazara briseis. Animal Conservation 13(2): 172 – 183.
↑
Kidner J., Theodorou P. et al. (2021): A brief history and popularity of methods and tools used to estimate micro-evolutionary forces. Ecology and Evolution
11(20): 13.723 – 13.743.
↑
Kinkler H., Löser S., Rehnelt K. (1987): 10 Jahre Erforschung des Moselapollofalters (Parnassius apollo vinningensis
Stichel 1899, Lepidoptera, Papilionidae) im modernen Weinbaugebiet der Mosel – ein Beitrag zu
seiner Rettung. Mitteilungen der Arbeitsgemeinschaft rheinisch-westfälischer Lepidopterologen 5: 74 – 96.
↑
Müller D., Hanisch K. (2020): Dramatischer Rückgang des Moselapollos Parnassius apollo vinningensis
Stichel, 1899 (Lep., Papilionidae). Melanargia 32(1): 1 – 8.
↑
Pecsenye K., Bereczki J. et al. (2007): Genetic differentiation among the Maculinea species (Lepidoptera:
Lycaenidae) in eastern Central Europe. Biological Journal of the Linnean Society 91(1): 11 – 21.
↑
Reed D.H., Frankham R. (2003):
Correlation between fitness and genetic diversity. Conservation Biology 17(1): 230 – 237.
↑
Reinhardt R., Harpke A. et al. (2020): Verbreitungsatlas der Tagfalter und Widderchen Deutschlands. Ulmer.
Stuttgart: 432 S.
↑
Schmidt A. (1997): Zur
aktuellen Situation des Mosel-Apollofalters Parnassius apollo vinningensis Stichel 1899 (Lep.
Papilionidae). Melanargia 9(2): 38 – 47.
↑
Schmitt T., Giessl A., Seitz A. (2003): Did Polyommatus icarus (Lepidoptera: Lycaenidae) have distinct
glacial refugia in southern Europe? Evidence from population genetics. Biological Journal of the Linnean Society 80(3):
529 – 538.
↑
Schmitt T., Habel J.C. et al. (2006): The Chalk-hill Blue Polyommatus coridon (Lycaenidae, Lepidoptera) in a highly fragmented landscape: How
sedentary is a sedentary butterfly? Journal of Insect Conservation 10(4): 311 – 316.
↑
Schmitt T., Hewitt G.M. (2004): The
genetic pattern of population threat and loss: A case study of butterflies. Molecular Ecology 13(1): 21 – 31.
↑
Schmitt T., Röber S., Seitz A. (2005): Is the last glaciation the only relevant event for the present genetic population structure of the
meadow brown butterfly Maniola jurtina (Lepidoptera: Nymphalidae)? Biological Journal of the Linnean Society 85(4):
419 – 431.
↑
Schmitt T., Seitz A. (2002): Influence of habitat fragmentation on the genetic structure of Polyommatus coridon (Lepidoptera:
Lycaenidae): Implications for conservation. Biological Conservation 107(3): 291 – 297.
↑
Sundberg M.D. (2010):
The tangled bank: An introduction to evolution. BioScience 60(3): 239 – 241.
↑
Tartally A., Thomas J. et al. (2019): Patterns of host use by brood parasitic Maculinea butterflies across Europe. Philosophical
Transactions of the Royal Society B 374(1.769): 20180202.
↑
Terrell K.A., Crosier A.E. et al. (2016): Continued decline in genetic diversity among wild cheetahs (Acinonyx jubatus) without further loss of
semen quality. Biological Conservation 200: 192 – 199.
↑
Újvári B., Madsen T. et al. (2002): Low genetic diversity threatens imminent extinction for the Hungarian meadow
viper (Vipera ursinii rakosiensis). Biological Conservation 105(1): 127 – 130.
↑
Újvári B., Olsson M., Madsen T. (2005): Discrepancy in mitochondrial and nuclear polymorphism in meadow vipers (Vipera ursinii) questions the
unambiguous use of mtDNA in conservation studies. Amphibia-Reptilia 26(3): 287 – 292.
↑
Vandewoestijne S., Nève G., Baguette M. (1999): Spatial and temporal population genetic structure of the butterfly Aglais urticae
L. (Lepidoptera, Nymphalidae). Molecular Ecology 8(9): 1.539 – 1.543.
↑
Vrba P., Grill S. et al. (2021): How do adults of the critically endangered hermit butterfly (Chazara
briseis) utilise their habitat? (Lepidoptera, Satyrinae). Journal of Insect Conservation 25(1): 39 – 48.
Fußnoten

