Heike Schneider, Peter Meyer,
Maria Aljes, Heike Culmsee, Marco Diers, Agnes Förster und Christoph Leuschner
Zusammenfassung
Die Nationale Strategie zur biologischen Vielfalt sieht in Deutschland eine natürliche Entwicklung auf 5 % der Waldfläche vor. Um
die Naturnähe von Wäldern besser definieren und bewerten zu können, wurden verschiedene Parameter aus den Bereichen Waldstruktur,
Artenvielfalt und Waldfunktionen in 16 Waldgebieten unterschiedlichen Alters und unterschiedlicher Baumartenzusammensetzung im
Norddeutschen Tiefland untersucht. Als Referenz für die Naturnähe der Waldstruktur reifer Waldentwicklungsstadien dienten Daten aus
ostslowakischen Buchen-Urwäldern. Anhand dieser Daten wurde ein Old-Growth-Indikator (OGI) als ein Maß für die Ähnlichkeit der
Waldstruktur mit reifen Waldentwicklungsphasen entwickelt. Die Urwälder zeigten eine deutlich größere Spanne der Waldstrukturdaten als
die untersuchten heimischen Waldbestände. Einige alte Laubwälder wiesen jedoch ähnliche Werte auf. Die Untersuchungen zeigten außerdem,
dass alte Laubwälder im Mittel etwa doppelt so viel Kohlenstoff in der oberirdischen Biomasse speicherten wie junge Kiefernforste.
Letztere wiesen jedoch deutlich höhere Kohlenstoffvorräte im Mineralboden auf. Weiterhin nahm die Gesamtartenzahl der Krautschicht in
alten Laubwäldern im Vergleich zu Kiefern(misch)wäldern ab. Die Zahl der auf geschlossene Wälder spezialisierten Arten sowie der an
Totholz gebundenen Käfer- und Pilzarten stieg dagegen mit zunehmender Ähnlichkeit der Waldstruktur mit reifen Waldentwicklungsphasen
an. Geeignete Referenzdaten sind essenziell für die Beurteilung der natürlichen Waldentwicklung ohne direkten menschlichen Einfluss.
Für die erfassten Biodiversitätskenngrößen fehlten diese jedoch. Dennoch können Daten zur Biodiversität unterstützend zum OGI in die
Naturnähebewertung von Wäldern eingehen.
Natürliche Waldentwicklung – Naturnähe – Waldstruktur – Biodiversität – KohlenstoffspeicherungAbstract
The German National Strategy on Biological Diversity aims for natural development on 5 % of the forest area. To better define and
assess the level of forest naturalness, various parameters of forest structure, species diversity and forest functions were
investigated in 16 forest areas differing in age and tree species composition in the North German Plain. Data collected in Slovakian
primeval beech forests served as a reference for naturalness of forest structure of old-growth forests. Using this data, an old-growth
indicator (OGI) was developed as a measure of similarity to old-growth forest structures. In general, these primeval forests were
characterised by a significantly wider range of values of forest-structure variables than the forest stands studied in northern German
lowlands. However, certain deciduous forests with management history also met some of the old-growth criteria which characterise
primeval forests. The study further found that old deciduous forests store on average about double the amount of carbon in the
aboveground biomass than the younger pine forests. Then again, carbon pools in the mineral soil were considerably higher under pine
forests. The total number of plant species decreased in old deciduous forests compared to (mixed) pine forests. In contrast, the number
of plant species dependent on closed forest canopies as well as the number of saproxylic species of beetles and fungi increased with
similarity to old-growth forest structures. Suitable references are essential when assessing natural forest development without direct
human impact. So far, such comparable references for biodiversity variables are absent. However, biodiversity information can indeed be
used in addition to the OGI indicator developed here when quantifying the naturalness of forests.
Natural forest development – Naturalness – Forest structure – Biodiversity – Carbon sequestrationInhalt
1 Einleitung
Vor dem Hintergrund klimatischer Veränderungen steht auch im Wald die Steigerung von Anpassungsfähigkeit und Resilienz an oberster
Stelle. Die Förderung von Vielfalt, Heterogenität und grundlegenden Waldfunktionen spielt dabei eine große Rolle. Neben Wiederbewaldung,
Waldumbau und naturnaher Waldbewirtschaftung gilt die Erhöhung des Anteils an Wäldern mit natürlicher Entwicklung als ein möglicher
Lösungsweg (Höltermann et al. 2019). Die Bundesregierung verfolgte mit der Nationalen
Strategie zur biologischen Vielfalt u. a. das Ziel einer natürlichen Entwicklung auf 5 % oder circa 5.500 km² der Waldfläche Deutschlands
bis zum Jahr 2020 (BMU 2007). Im Kontext einer naturnahen Waldbewirtschaftung bzw. einer
natürlichen Waldentwicklung stellt sich die grundlegende Frage, was eigentlich unter den Begriffen „natürlich“ und „naturnah“ genau zu
verstehen ist und wie dies bewertet werden kann. Kowarik (1999) unterscheidet zwischen
„Naturnähe“ als Maß für die Ursprünglichkeit von Standorten oder Vegetationsausschnitten aus einer historischen Perspektive und
„Hemerobie“ als Maß für den menschlichen Kultureinfluss auf Grundlage der heutigen standörtlichen Verhältnisse. Überlässt man Wälder der
natürlichen Entwicklung, so wird angenommen, dass durch nachlassende menschliche Einflussnahme der Grad der Hemerobie sinkt und in vielen
Fällen die Naturnähe im Gegenzug ansteigt. Bislang fehlen Informationen darüber, wie genau und wie schnell diese Entwicklung abläuft.
Bestehende Bewertungsmethoden verzichten auf eine Validierung der Naturnähebewertung anhand von Referenzwerten aus Urwäldern, die hier als
natürlich entstandene Wälder mit einheimischen Baumarten und ohne Anzeichen für menschliche Aktivitäten oder gestörte ökologische Prozesse
verstanden werden (Wirth et al. 2009; Winter et al.
2010; Schall, Ammer 2013; Kahl, Bauhus
2014; FAO 2020).
Aufgrund der langen, intensiven Nutzungsgeschichte unserer Landschaft fehlen Urwälder hierzulande vollständig und trotz
der Zunahme von Wäldern mit natürlicher Entwicklung (Engel 2019) machen längerfristig
ungenutzte Wälder nur einen Bruchteil der Waldfläche Deutschlands aus (Sabatini et al. 2018).
Die Erhebung vergleichbarer Referenzdaten für die Naturnähebewertung heimischer Wälder ist daher sehr schwierig. Eine Möglichkeit, die
Kennzeichen naturnaher Wälder zu quantifizieren, besteht darin, die verbliebenen Urwaldrelikte in Naturräumen des östlichen Mitteleuropas,
die den naturräumlichen Verhältnissen in Deutschland ähneln, zu nutzen. Allerdings bilden diese Restbestände nur einen Ausschnitt der
gesamten natürlichen Waldentwicklung ab. Hier werden v. a. reife Waldentwicklungsstadien naturnaher Wälder betrachtet, während frühe
Sukzessions- und Entwicklungsstadien, die sich nach größeren Störungen einstellen, weitgehend fehlen (Swanson et al. 2011). Durch Untersuchungen in Urwaldrelikten ist das sog. „Old-Growth-Stadium“ von Wäldern in der Literatur
verhältnismäßig klar definiert (Bauhus et al. 2009; Wirth
et al. 2009). Die Bedeutung und die Auswirkungen großflächiger Störungen können allerdings nur erahnt werden.
Für das Projekt „Wildnis Naturerbe“ (WiNat) untersuchten wir bewirtschaftete sowie aus der Nutzung entlassene Waldbestände im
Norddeutschen Tiefland mit dem Ziel, geeignete Kenngrößen zur Naturnähebewertung zu identifizieren und den Grad der Naturnähe zwischen den
Wäldern zu vergleichen. Dazu wurden verschiedene Kenngrößen aus den Bereichen Waldstruktur, Waldfunktionen und Biodiversität erhoben und
der Einfluss der Nutzungsaufgabe untersucht. Als Referenz maximaler Naturnähe konnten wir für die Waldstrukturdaten auf vergleichbare
Untersuchungen aus ostslowakischen Buchen-Urwäldern zurückgreifen (Feldmann 2018; Glatthorn 2018). Im Folgenden werden die verwendeten Erfassungsmethoden sowie die Ergebnisse
erläutert und diskutiert. Insbesondere wird der anhand unserer Daten und der Urwald-Referenzdaten entwickelte Waldstruktur-Indikator
(Old-Growth-Indikator – OGI) dargestellt, der – unterstützt durch weitere Daten zur Artenvielfalt und Kohlenstoffspeicherung –
eine Naturnähebewertung der Waldbestände ermöglicht.
2 Material und Methoden
2.1 Untersuchungsgebiete
Die 16 Untersuchungsgebiete verteilen sich über das gesamte Norddeutsche Tiefland und repräsentieren die auf den Standorten
großflächig verbreiteten potenziellen natürlichen Waldgesellschaften (Abb. 1, Tab. A im unter
Online-Zusatzmaterial). Dabei handelt es sich
vorwiegend um basenarme bis basenreichere Buchenwälder sowie im Fall der drei Untersuchungsgebiete Weißhaus, Prösa und Zschornoer Wald im
Ostdeutschen Tiefland um Eichen-Kiefernmischwälder auf Sandböden. Die Untersuchungsgebiete sind entweder aus der Nutzung entlassene
Naturwaldreservate bzw. bewirtschaftete Bestände der Landesforsten Niedersachsen und Sachsen-Anhalt oder Teil des Nationalen Naturerbes im
Eigentum der Naturerbe GmbH, Tochter der Deutschen Bundesstiftung Umwelt (DBU).
In jedem Untersuchungsgebiet haben wir drei Waldbestände in Anlehnung an die Waldkategorien der DBU Naturerbe GmbH
ausgewählt (DBU Naturerbe GmbH 2014; Tab. 1).
Wälder mit natürlicher Entwicklung (Kategorie N) sind charakterisiert durch über 100-jährige Buchen- bzw. Eichenbestände, in denen
die Bewirtschaftung vor bis zu 40 Jahren eingestellt wurde und die sich seitdem in natürlicher Entwicklung befinden. In der näheren
Umgebung dieser Bestände wurden jeweils folgende Wälder mit ähnlichen Standortbedingungen ausgewählt: ein Kiefern-Laubmischwald
(> 80 Jahre) als Beispiel für einen Wald, der kurzfristig in eine natürliche Entwicklung überführt wird (Kategorie ÜK), und ein
bewirtschafteter Kiefernwald (< 80 Jahre) als Beispiel für einen Wald, der langfristig in eine natürliche Entwicklung überführt wird
(Kategorie ÜL). Für die Datenerhebungen wurden in jedem Waldbestand zehn zufällig verteilte Probekreise mit einem Mindestabstand
von 66 m zueinander und mit einer Fläche von jeweils 1.000 m² eingerichtet.
|
Waldkategorie
|
Bestandsbeschreibung
|
Bewirtschaftung
|
N
| Natürliche Entwicklung | Wälder älter als 100 Jahre, Baumartenzusammensetzung ähnelt der potenziellen natürlichen Vegetation | Wälder befinden sich in natürlicher Entwicklung ohne weitere Eingriffe |
ÜK
| Überführung kurzfristig | Mischbestände aus Kiefern älter als 80 Jahre und standortheimischen Laubbaumarten im Hauptbestand | Begrenzte Eingriffe zur Entwicklungssteuerung in ≤ 20 Jahren, nicht standortheimische Baumarten werden sukzessive
entnommen |
ÜL
| Überführung langfristig | Kiefernbestände jünger als 80 Jahre, standortheimische Laubbaumarten kommen nur im Unterstand vor | Langfristige Entwicklungssteuerung in > 20 Jahren, nicht standortheimische Baumarten werden sukzessive
entnommen |
OG
| Old-Growth-Referenz (slowakische Buchen-Urwälder) | Buchen-Urwälder, ca. 400 Jahre | Keine bekannte forstliche Bewirtschaftung |
Tab. 1: Beschreibung der Kategorien der Untersuchungsbestände in Anlehnung an die Waldkategorien der DBU Naturerbe GmbH (DBU 2014) und Beschreibung der Kategorie der Referenzbestände (slowakische
Buchen-Urwälder).
Table 1: Description of the categories of the studied forest stands according to forest categories of DBU Natural Heritage
(DBU 2014) and description of the category of reference stands (Slovakian primeval
beech forests).
Als Referenzbestände maximaler Naturnähe dienten die drei ungefähr 400 Jahre alten ostslowakische Buchen-Urwälder Kyjov, Havešová,
Stužica (Kategorie OG = Old-Growth-Referenz; Tab. 1), aus denen uns Waldstrukturdaten
zur Verfügung gestellt wurden, die mit derselben Methodik erfasst wurden (Feldmann 2018;
Glatthorn 2018; Abb. 1). In jedem der drei
Referenzbestände wurden zwölf Probekreise erfasst. Trotz gewisser Unterschiede in den edaphischen (den Boden betreffenden) und
klimatischen Bedingungen zwischen den Untersuchungs- und Referenzbeständen ist davon auszugehen, dass die Wuchsbedingungen und die
Strukturdynamik innerhalb des natürlichen Verbreitungsgebiets der mitteleuropäischen Rotbuchenwälder hinreichend ähnlich sind (Markgraf 1931; Peters 1992).
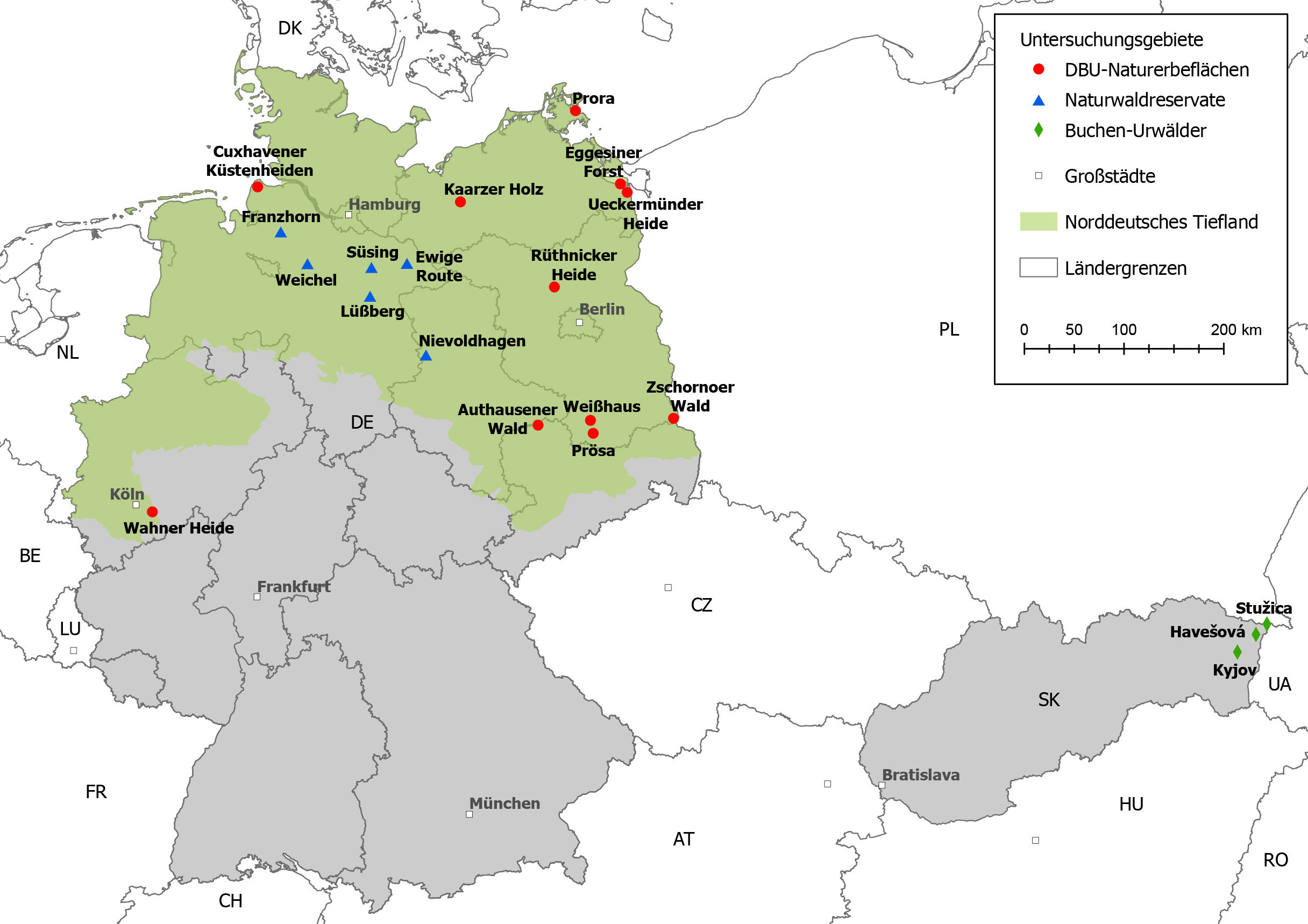
Abb. 1: Lage der Untersuchungsgebiete im Norddeutschen Tiefland und Lage der als Referenz für die Waldstrukturdaten dienenden
slowakischen Buchen-Urwälder. Weitere Informationen zu den Waldbeständen in Tab. A im
Online-Zusatzmaterial.
Fig. 1: Location of studied forest areas in the North German Plain and location of the Slovakian primeval beech forests as
reference for forest-structure data. For further information on the forest stands, see Table A in the
online supplementary material.
2.2 Erfassungs- und Auswertungsmethoden
In allen Waldbeständen im Norddeutschen Tiefland untersuchten wir verschiedene Parameter aus den Bereichen Waldstruktur, Waldfunktion
und Biodiversität. Die Waldstruktur erfassten wir nach der Methodik von Meyer et al. (2013).
Dabei wurden der lebende Derbholzbestand ≥ 7 cm Brusthöhendurchmesser (BHD = Durchmesser eines Baumes in 1,3 m Höhe), Totholz (stehendes
Totholz ≥ 7 cm BHD, liegendes Totholz ≥ 20 cm Durchmesser am stärkeren Ende), die Baumartenverjüngung und Mikrohabitate innerhalb der
Probekreise erfasst.
Als funktionale Kenngrößen untersuchten wir die Kohlenstoffspeicherung und -sequestrierung (Kohlenstoffbindung). Die
Kohlenstoffvorräte in der oberirdischen Biomasse berechneten wir anhand der auf allen Probekreisen erfassten Waldstrukturdaten. Die
oberirdische Nettoprimärproduktion als Summe der Holz- und Streuproduktion erfassten wir auf sechs zufällig ausgewählten Probekreisen in
den N- und ÜL-Beständen unter Verwendung von Zuwachsmaßbändern und Streufängern in drei aufeinanderfolgenden Jahren (Methodenbeschreibung
in Förster et al. 2021a). Die Feinwurzelbiomasse und Feinwurzelnekromasse (organische Auflage
und Mineralboden 0 – 20 cm) und das Feinwurzelwachstum (0 – 20 cm Boden inklusive organischer Auflage) wurden auf denselben sechs
Probekreisen bestimmt (Förster et al. 2021b). Zur Messung des Feinwurzelwachstums nutzten wir
die Ingrowth-Core-Methode (Persson 1980; Majdi
1996). Zur Umrechnung von Biomasse in Kohlenstoff wurde ein Kohlenstoffgehalt von 50 % angenommen. Zusätzlich entnahmen wir
Bodenproben bis zu einer Tiefe von 60 cm in jedem Probekreis zur Ermittlung der Bodenkohlenstoffvorräte (Diers et al. 2021). Sämtliche erhobenen Kenngrößen aus den bewirtschafteten und den aus der Nutzung
entlassenen Wäldern wurden miteinander verglichen und Einflussfaktoren untersucht.
Als Biodiversitätskenngrößen wurden die Bodenvegetation, epiphytische Moose und Flechten, xylobionte Käfer (Holzkäfer) sowie Pilze
erfasst. Die Bodenvegetation (Krautschicht, bodenbewohnende Moose und Flechten, Verjüngung der Gehölze < 7 cm BHD) untersuchten wir in
jedem Probekreis auf einer Unterfläche von 100 m². In der Hauptvegetationsperiode von Mai bis Juli wurden für jede auftretende Art
prozentuale Deckungsgrade und die Höhe getrennt nach Schichten angegeben. Die Diversität epiphytischer Flechten und Moose erfassten wir
innerhalb der Probekreise auf allen Bäumen innerhalb einer Fläche von 500 m² (stehende Bäume bis zu einer Höhe von 2 m, liegendes Totholz
auf der ganzen Oberfläche). Für die Inventur xylobionter Käfer (inklusive fakultativ xylobionter Käfer, Altholzkäfer) installierten wir
eine Flugfensterfalle pro Waldbestand, die monatlich von Mai bis September/Oktober geleert wurde. Die Pilzinventuren führten wir auf vier
zufällig ausgewählten Probekreisen pro Waldbestand durch. Mittels einer zeitstandardisierten Begehung wurde jeder Probekreis jeweils
45 min pro Person und Erfassungsdurchgang begangen und nach den relevanten Pilzvorkommen hin visuell untersucht. Zusätzlich wurde der
Gesamtbestand auf der Suche nach naturschutzfachlich relevanten bzw. gebietstypischen Arten abgegangen. In drei Erfassungsdurchgängen pro
Jahr kartierten wir den Frühjahrs-, Haupt- und Spätherbstaspekt folgender Artengruppen: Ektomykorrhiza-Pilze, Holz- und Streuzersetzer
(Basidiomycota: Agaricales, Boletales, Cantharellales, Russulales, Thelephorales und polyporoide sowie corticioide Pilze; alle Ascomyzeten
mit großen Fruchtkörpern).
Aus den von uns erhobenen Waldstrukturdaten und Urwald-Referenzdaten konnte der OGI als Instrument für die Naturnähebewertung der
Waldstruktur entwickelt werden. Der OGI bezieht sich ausschließlich auf die Waldstruktur reifer Waldentwicklungsstadien. Für eine
umfassende Naturnähebewertung, die auch größere Störungs- und Sukzessionsflächen berücksichtigt (Swanson et al. 2011), fehlen belastbare
Daten über deren natürliche Häufigkeit und Flächenausdehnung in mitteleuropäischen Wäldern. Für den OGI identifizierten wir
27 urwaldtypische Kenngrößen aus 10 thematischen Gruppen (Totholz, Dichte Derbholz, Dichte Verjüngung, Waldentwicklungsphase, Durchmesser,
Habitate, Zersetzungsgrad, Autochthonie, Baumarten, sukzessionaler Status) aus den vorliegenden Waldstrukturdaten der Buchen-Urwälder und
verglichen diese mit unseren Untersuchungsbeständen. Eine detaillierte Methodenbeschreibung zur Entwicklung des OGI findet sich
in Kasten 1 und in Meyer et al. (2021). Anhand
einer Hauptkomponentenanalyse (PCA) der 27 OGI-Kenngrößen wurden Unterschiede in den Wertespannen zwischen Untersuchungs- und
Referenzbeständen deutlich gemacht.
Kasten 1: Herleitung des Old-Growth-Indikators (OGI) zur Naturnähebewertung der Waldstruktur.
Box 1: Generating the old-growth indicator (OGI) for the assessment of naturalness of forest structure.
Mit dem OGI sollte eine Methode entwickelt werden, um die Ähnlichkeit genutzter bzw. aus der Nutzung genommener Wälder mit
Old-Growth-Stadien von Buchen-Urwäldern anhand ausgewählter, charakteristischer Strukturmerkmale zu bewerten. Als Referenz nutzten wir
3 ostslowakische Buchen-Urwälder, in denen im Rahmen von Doktorarbeiten auf jeweils 12 Plots Waldstrukturaufnahmen nach Meyer et al. (2013) durchgeführt wurden. Für die Entwicklung des OGI berechneten wir aus diesen
Daten zunächst für jeden Plot 134 Strukturvariablen (z. B. prozentualer Anteil von Bäumen einer bestimmten Durchmesserklasse,
Totholzvolumen des liegenden und stehenden Totholzes verschiedener Durchmesserklassen, Anzahl verschiedener Baumarten im Derbholz).
Dieses Set an Variablen wurde in einem nächsten Schritt von den Autorinnen und Autoren gutachterlich geprüft und auf eine Auswahl von
41 Variablen reduziert, die für die Beschreibung der Waldstruktur relevant und möglichst einfach zu erheben und zu interpretieren sind.
Anschließend prüften wir mit einem nicht-parametrischen Kruskal-Wallis-Rangsummentest (p ≤ 0,05), für welche der 41 ausgewählten
Variablen keine signifikanten Unterschiede zwischen den 3 Buchen-Urwäldern vorlagen. Diese wurden als „urwaldtypisch“ betrachtet und in
10 thematische Gruppen eingeordnet, die nach unserer Auffassung essenziell für die Beschreibung der Waldstruktur reifer Buchenwälder
sind (Tab. B im Online-Zusatzmaterial).
Um miteinander korrelierende Variablen auszuschließen, führten wir anschließend eine Überprüfung der Multikollinearität innerhalb
der thematischen Gruppen durch (prog-reg-Protokoll in SAS 9.4®, option vif collin). Dabei wurde ein variance inflation
factor von > 10 und/oder tolerance-Wert von < 0,2 als Schwellenwert verwendet, um die Variablenselektion vergleichbar zu halten.
Für Variablen, bei denen eine Multikollinearität nachgewiesen werden konnte, wurde jeweils die Variable behalten, die in
Strukturbeschreibungen in der Literatur weiter verbreitet und/oder leichter zu berechnen ist. Daraus ergab sich ein Set aus
27 Variablen für die Berechnung des OGI. Im Folgenden wird die Zuordnung der Variablen zu den jeweiligen Gruppen sowie deren Berechnung
beschrieben:
● Sukzessionaler Status: Old-Growth-Stadien sind insbesondere durch die Dominanz spätsukzessionaler Baumarten
gekennzeichnet (Korpel' 1995). Der momentane sukzessionale Status des
Derbholzbestands (Brusthöhendurchmesser – BHD ≥ 7 cm) sowie der Verjüngung wurde ermittelt, indem der Anteil der Stammzahl von
Pionierbaumarten (z. B. Birke – Betula, Pappel – Populus, Kiefer – Pinus) mit dem Faktor 1, der Anteil
der Stammzahl von Intermediärbaumarten (z. B. Eiche – Quercus, Ahorn – Acer, Esche – Fraxinus) mit dem
Faktor 2 und der Anteil der Anzahl schattentoleranter Klimaxbaumarten (Buche – Fagus) mit dem Faktor 3 multipliziert
wurde. Die Summe der Werte ergibt den sukzessionalen Status. ● Waldentwicklungsphase: Für die Berechnung der Waldentwicklungsphase eines Plots wurden zunächst alle Bäume
des Derbholzbestands anhand ihres BHD in sog. „natürliche Altersklassen“ eingeteilt (NFP
2001) mit der Abstufung 1 (< 20 cm), 2 (20 – 35 cm), 3 (> 35 – 50 cm), 4 (> 50 – 80 cm) und 5
(> 80 cm). Auch hier wurde aus den Anteilen der Stammzahlen pro Klasse die Summe gebildet, um die Variable zu erhalten. Die
zweite Variable dieser thematischen Gruppe ist die Anzahl der vorhandenen Altersklassenstufen. ● Baumartendiversität: Alte Buchenwaldstadien weisen eine sehr geringe Baumartendiversität auf (Leuschner 2015). Diesem Merkmal wurde mit der Variablen „Baumartenanzahl“
Rechnung getragen. ● Autochthone Baumarten: Die Baumartenzusammensetzung in Old-Growth-Beständen besteht ausschließlich aus
autochthonen Gehölzen. Daher wurde der Anteil nicht-autochthoner Baumarten sowohl im Derbholzbestand als auch in der
Verjüngung als Indikator für anthropogene Einflüsse auf die Bestände gewertet. ● Differenzierung der Baumdimensionen: Als Annäherung an die für Urwälder als typisch beschriebene umgekehrte
Sigmoidkurve der Durchmesserverteilung (Goff, West 1975; Westphal et al. 2006; Feldmann et al.
2018) verwendeten wir den minimalen und maximalen BHD, die BHD-Spanne und die Anzahl von Bäumen mit
einem Durchmesser von mehr als 80 cm („Urwaldriesen“). ● Dichte: Hier wurden sowohl Anzahl der Stämme als auch Volumen des lebenden Derbholzbestands pro ha
ausgewählt. ● Verjüngungsdichte: Zur Beschreibung der Verjüngung in Old-Growth-Beständen wurden die Merkmale Gesamtdichte der
Verjüngung (Stückzahl pro ha) sowie die Dichte innerhalb der Höhenklasse 0,5 – 1,5 m und ≥ 1,5 m
ausgewählt. ● Totholz: Durch die natürliche Alterung von Old-Growth-Beständen kommt es zu einer starken Akkumulation von Totholz
(Bauhus et al. 2009). Diese Gruppe wird von 6 Variablen gebildet:
Gesamttotholzvolumen sowie das einzeln berechnete Volumen stehenden und liegenden Totholzes in 3 bzw.
2 Durchmesserklassen. Die Old-Growth-Bestände stechen insbesondere in Bezug auf starkes stehendes Totholz hervor,
wohingegen Totholz in den geringeren Durchmesserklassen aufgrund einer geringen Selbstdurchforstungsrate weniger vertreten
ist. ● Zersetzungsgrad: Totholz durchläuft während der Zersetzung verschiedene Stadien, die in die Zersetzungsgrade 1
(frisch abgestorben) bis 5 (stark zersetzt, vermodert) (Meyer et al. 2013)
eingeordnet werden. Die relative Häufigkeit der Anzahl der Objekte im jeweiligen Stadium wurde mit dem Zersetzungsgrad
multipliziert und die Summe ergibt den mittleren Zersetzungsgrad. Typischerweise sind Old-Growth-Bestände durch einen
hohen Wert gekennzeichnet, da zum Erreichen eines hohen Zersetzungsgrads wesentlich mehr Zeit benötigt wird als für niedrige
Zersetzungsgrade (Müller-Using, Bartsch 2009). ● Mikrohabitate: Es wurden drei Typen von Mikrohabitaten gewählt, die wichtige Merkmale für Old-Growth-Bestände
darstellen: Die Anzahl von Höhlenbäumen und Bäumen mit Pilzkonsolen ist in Old-Growth-Beständen hoch, während
die Anzahl von Wurzeltellern durch eine starke Schwankungsbreite gekennzeichnet ist.
Für das Bootstrapping nach der Variablenselektion siehe Abschnitt 1 im Online-Zusatzmaterial.
Insgesamt identifizierten wir 11 verschiedene Möglichkeiten der Übereinstimmung zwischen kontinuierlichen oder
Punktvariablenwerten, bei teilweiser oder vollständiger Übereinstimmung der Schwankungsbereiche (Abb. A im Online-Zusatzmaterial). Die Erklärung zu den einzelnen
Fallberechnungen findet sich in Tab. C im Online-Zusatzmaterial.
Die statistische Datenanalyse führten wir mit den Programmen R (Version 3.5.1, R Core Team, 2018) und SAS 9.4® durch.
Weitere Einzelheiten können außerdem der „Methodenbeschreibung Naturnähe Monitoring“ sowie dem Abschlussbericht des Projekts „WiNat –
Wildnis Naturerbe“ entnommen werden (Culmsee et al. 2021).
3 Ergebnisse
3.1 Waldstruktur
Baumartenzusammensetzung, Bestandsalter und nutzungsfreie Zeit in den drei Waldkategorien N, ÜK und ÜL wirkten sich deutlich auf die
Bestandsstruktur aus. Die Werte für Derbholz- und Totholzvolumen, Durchmesserspanne und Vorkommen von Mikrohabitaten (Höhlenbäume)
unterschieden sich signifikant zwischen den untersuchten Waldkategorien (Tab. 2). Auch wenn
Laubwälder der Kategorie N deutlich höhere Werte aufwiesen als die Kiefernwälder (Kategorie ÜL), so zeigte der Vergleich mit
Urwäldern noch einmal deutliche Unterschiede. Das Totholzvolumen war in den Buchen-Urwäldern (Kategorie OG) fast zehnmal so hoch wie in
den Wäldern der Kategorie N, die mittlere Anzahl an Totholzstämmen, Höhlenbäumen und Verjüngungspflanzen immerhin dreimal höher. Die
Ergebnisse der PCA zeigten, dass Wälder mit natürlicher Entwicklung insgesamt eine größere Spanne der 27 für den OGI verwendeten
Waldstrukturdaten (Tab. B im Online-Zusatzmaterial)
aufwiesen als die Kiefern(misch)wälder (Abb. 2). Die Wertespannen der Wälder mit natürlicher
Entwicklung wichen trotz gewisser Überlappungen deutlich von den Buchen-Urwäldern ab. Es zeigte sich auch, dass der OGI nach einer
Nutzungsaufgabe im Laufe der Zeit zunimmt (Abb. 3).
|
Gruppe
|
ÜL
|
ÜK
|
N
|
OG
|
Mittleres Derbholzvolumen gesamt [m³ je ha] | 289a | 312a | 455b | 591c |
Mittlere Durchmesserspanne Derbholz [cm] | 33a | 44b | 56c | 74d |
Mittleres Totholzvolumen gesamt [m³ je ha] | 6a | 12b | 16b | 153c |
Mittlere Anzahl Totholzstämme [je ha] | 48a | 61a | 38b | 111c |
Mittlere Anzahl Höhlenbäume [je ha] | 2a | 5b | 8c | 24d |
Mittlere Anzahl Verjüngungspflanzen [je ha] | 7.160a | 17.127a | 11.381a | 33.252b |
Gesamtartenzahl Pilze | 846 | 950 | 1.033 | K. A. |
Gesamtartenzahl Holzpilze (Xylobionte) | 469 | 516 | 538 | K. A. |
| 16 | 15 | 25 | K. A. |
Gesamtartenzahl Käfer | 476 | 510 | 547 | K. A. |
Gesamtartenzahl Holzkäfer (Xylobionte) | 200 | 330 | 349 | K. A. |
| 4 | 4 | 2 | K. A. |
Gesamtartenzahl krautige Arten (Bodenvegetation) | 110 | 110 | 98 | K. A. |
Gesamtartenzahl Gehölzarten (Bodenvegetation) | 29 | 29 | 25 | K. A. |
Gesamtartenzahl Moose/Flechten (Bodenvegetation) | 25 | 26 | 18 | K. A. |
| 37 | 43 | 45 | K. A. |
Gesamtartenzahl epiphytischer Moose | 41 | 41 | 43 | K. A. |
Gesamtartenzahl epiphytischer Flechten | 17 | 22 | 28 | K. A. |
N = natürliche Entwicklung, ÜK = Überführung kurzfristig, ÜL = Überführung langfristig, OG = Old-Growth-Referenz,
K. A. = keine Angabe |
Tab. 2: Mittelwerte verschiedener Waldstrukturparameter und Gesamtartenzahlen der Pilz-, Käfer- und Pflanzendiversität in allen
Untersuchungsbeständen der drei Waldkategorien N, ÜK und ÜL sowie in den Buchen-Urwäldern (Kategorie OG). Für weitere
Informationen zu den Waldkategorien siehe Tab. 1. Unterschiedliche Buchstaben a, b,
c, d zeigen signifikante Unterschiede zwischen den Mittelwerten der Waldkategorien (Wilcoxon-Rangsummentest mit p ≤ 0,05; Daten
der Buchen-Urwälder aus Feldmann 2018 und Glatthorn 2018).
Table 2: Mean values of forest-structure parameters and absolute species numbers of fungal, beetle and plant diversity in all
studied stands of the three forest categories N, ÜK and ÜL as well as in the Slovakian primeval beech forests (category OG). For
more information on the forest categories, see Table 1. Different letters a, b, c, d
indicate significant differences between the mean values of forest categories (Wilcoxon rank sum test with p ≤ 0.05; data on
primeval beech forests from Feldmann 2018 and Glatthorn 2018).
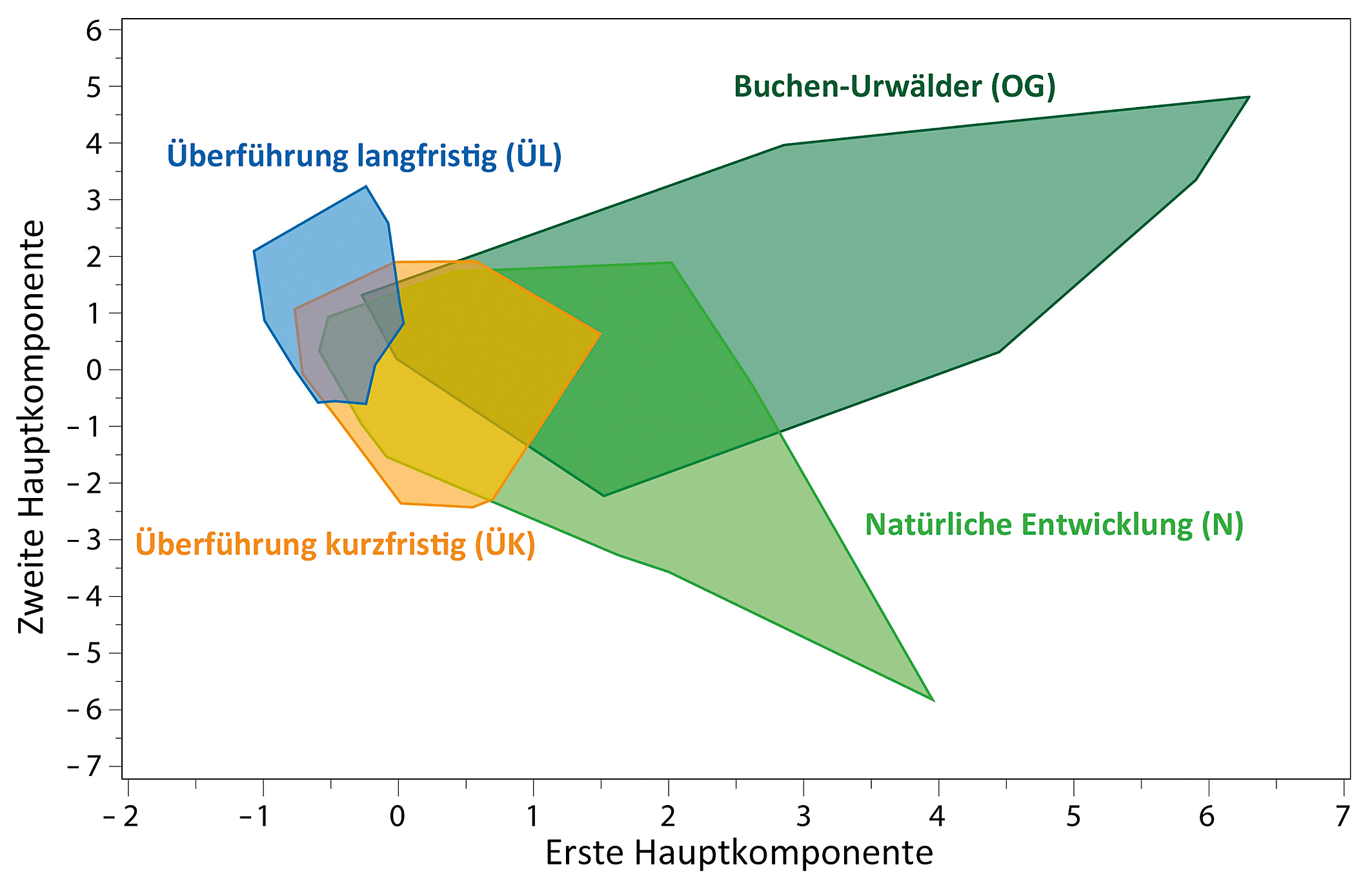
Abb. 2: Hauptkomponentenanalyse (PCA) zum Vergleich der Wertespanne der Waldstrukturdaten auf Plotebene zwischen den slowakischen
Buchen-Urwäldern (Kategorie OG = Old-Growth-Referenz; n = 36) und den untersuchten Waldbeständen (Kategorien N = natürliche
Entwicklung, ÜK = Überführung kurzfristig und ÜL = Überführung langfristig; jeweils mit n = 160). Für weitere Informationen zu
den Waldkategorien siehe Tab. 1.
Fig. 2: Principal component analysis (PCA) for comparison of range of forest-structure data between Slovakian primeval beech
forests (category OG = old-growth reference; n = 36) and the forest stands studied (categories N = natural development,
ÜK = transfer in the short term, ÜL = transfer in the long term; with n = 160 each). For more information on the forest categories,
see
Table 1.
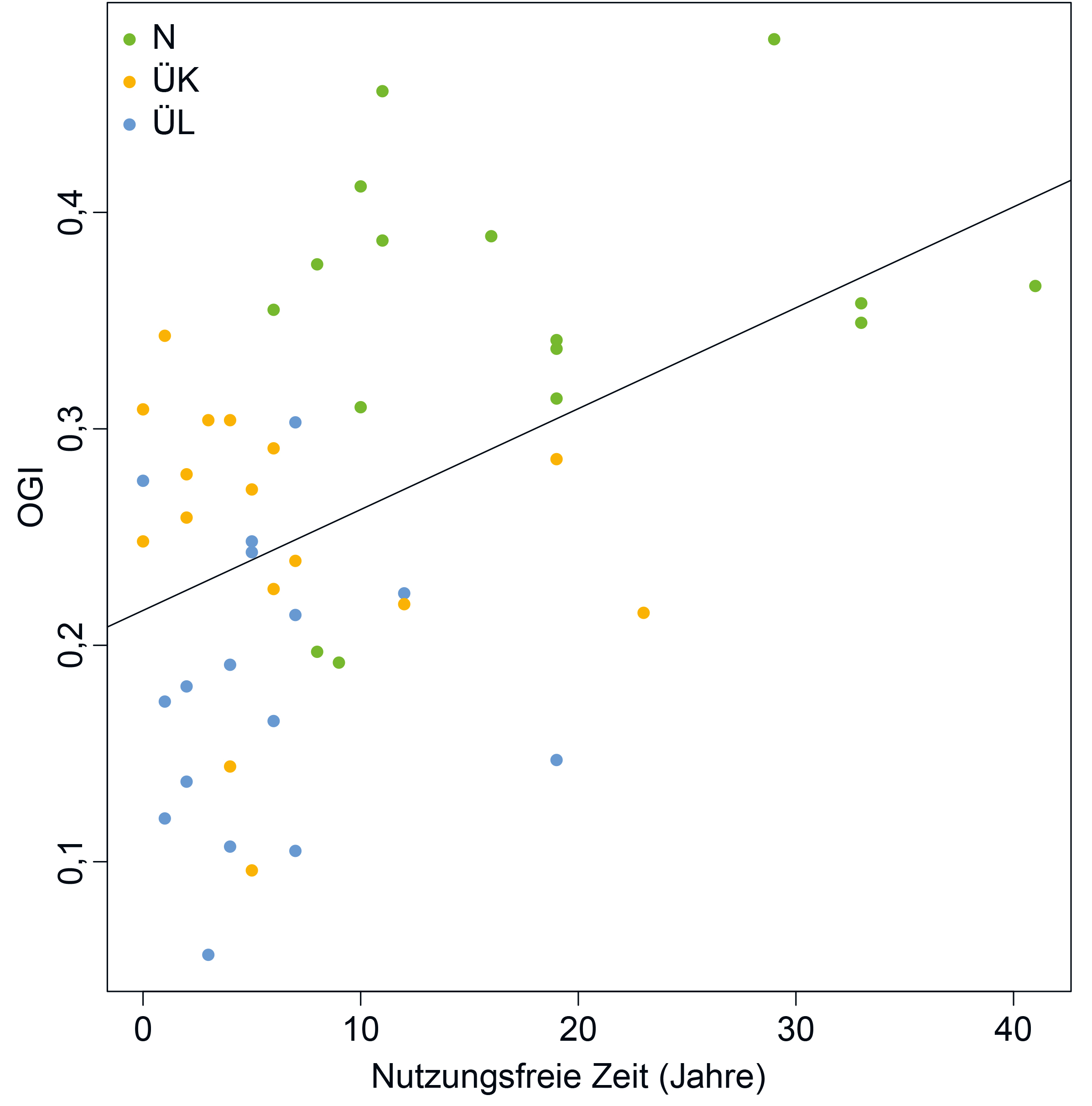
Abb. 3: Korrelation zwischen dem Old-Growth-Indikator (OGI) und der nutzungsfreien Zeit dargestellt für Bestände der drei
Waldkategorien N = natürliche Entwicklung, ÜK = Überführung kurzfristig, ÜL = Überführung langfristig. Korrelationskoeffizient
(Pearson) für alle drei Waldkategorien = 0,45 mit p = 0,001. Für weitere Informationen zu den Waldkategorien siehe Tab. 1.
Fig. 3: Correlation between old-growth indicator (OGI) and the time since last silvicultural intervention for forest stands of
the three categories N = natural development, ÜK = transfer in the short term, ÜL = transfer in the long term. Correlation
coefficient (Pearson) for all three forest categories = 0.45 with p = 0.001. For more information on the forest categories, see
Table 1.
Demnach ergaben sich insgesamt signifikant höhere OGI-Werte für Buchen-Urwälder im Vergleich zu den heimischen Untersuchungsbeständen
aller drei Kategorien (Abb. 4; Meyer et al.
2021). Die Mittelwerte des OGI für alle Bestände jeweils einer Waldkategorie betrugen 0,83 für Buchen-Urwälder (Kategorie OG),
0,35 für Wälder der Kategorie N gefolgt von 0,26 für Wälder der Kategorie ÜK und 0,18 für Wälder der Kategorie ÜL. Das Vorhandensein
autochthoner Baumarten und Mikrohabitate hatten in den meisten heimischen Untersuchungsbeständen (Kategorien N, ÜK, ÜL) einen hohen
Einfluss auf den OGI mit 27 % bzw. 17 % (übrige Waldstrukturparameter jeweils ca. 7 %). Die Kategorie ÜK unterschied sich von ÜL v. a. in
den Gruppen „Waldentwicklungsphase“ und „Totholz“. Der im Vergleich zu den Kategorien ÜK und ÜL höhere Gesamtwert des OGI in Wäldern der
Kategorie N basierte v. a. auf hohen Werten in den Gruppen „sukzessionaler Status“ und „Anzahl Baumarten“.
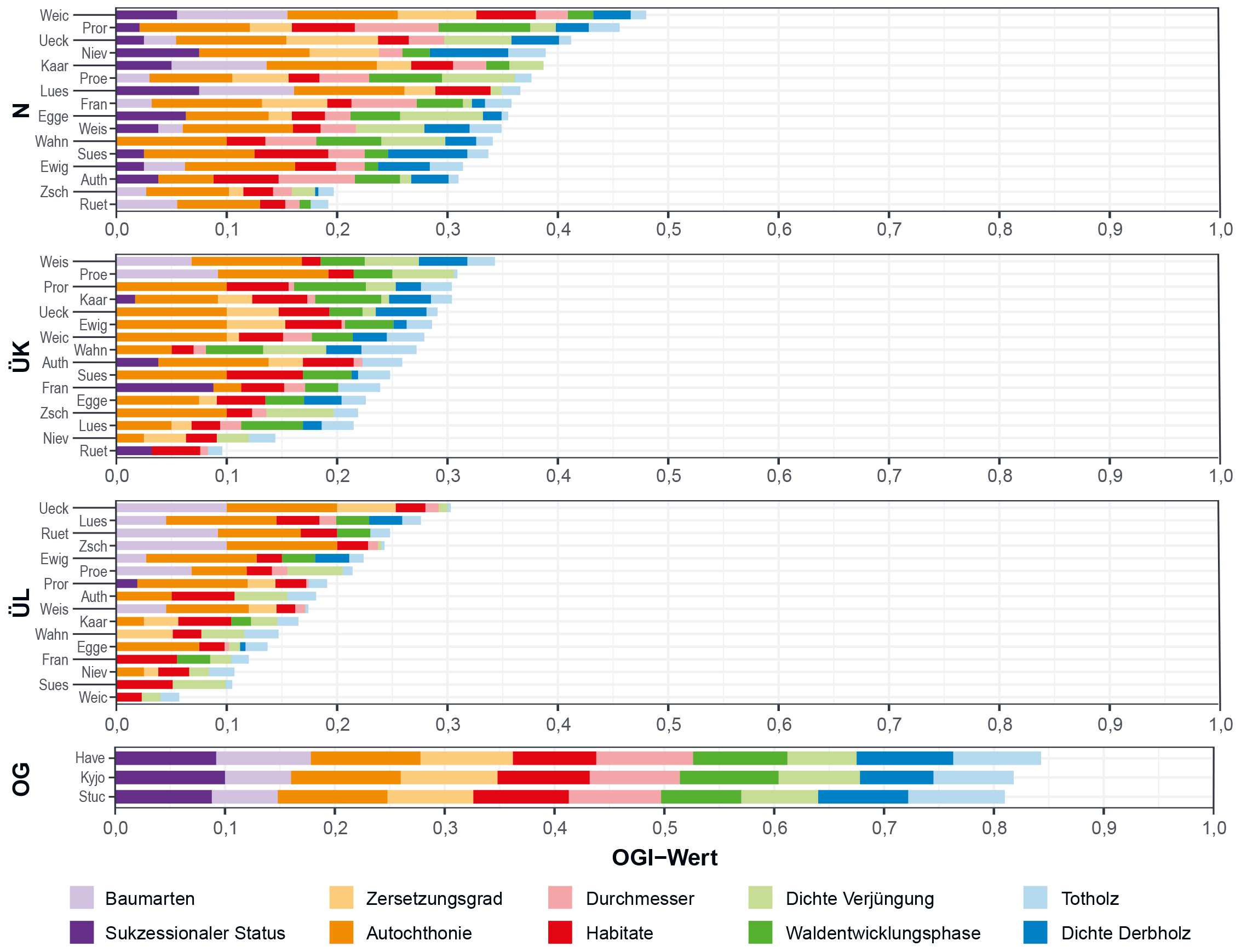
Abb. 4: Einzelwerte und aggregierte Werte des Old-Growth-Indikators (OGI) für die Untersuchungsgebiete der drei Waldkategorien
N = natürliche Entwicklung, ÜK = Überführung kurzfristig und ÜL = Überführung langfristig und der slowakischen Buchen-Urwälder
(Kategorie OG = Old-Growth-Referenz) mit jeweils gleicher Gewichtung der zehn thematischen Gruppen. Jede der Waldkategorien N, ÜK
und ÜL beinhaltet 16 Untersuchungsbestände; in den Buchen-Urwäldern wurden 3 Waldbestände untersucht (Abkürzungen der
Untersuchungsgebiete und weitere Bestandsinformationen siehe Tab. A im Online-Zusatzmaterial). Die Entwicklung des OGI
und der dafür genutzten Gruppen urwaldtypischer Kenngrößen ist in Kasten 1 näher
beschrieben. Für weitere Informationen zu den Waldkategorien siehe Tab. 1.
Fig. 4: Single and aggregated values of old-growth indicator (OGI) for the study areas of the forest categories N = natural
development, ÜK = transfer in the short term, ÜL = transfer in the long term and Slovakian primeval beech forests (category
OG = old-growth reference) with equal weight of the ten thematic groups. Each of the forest categories N, ÜK and ÜL comprises
16 study areas; in the primeval beech forests, three forest stands were investigated (abbreviations of the study areas and further
information on forest stands in Table A in the
online supplementary material). The development of the OGI and the utilised groups of typical old-growth attributes are explained
in
Box 1. For more information on the forest categories, see
Table 1.
3.2 Kohlenstoffspeicherung
Die hier untersuchten alten Laubwälder der Kategorie N speicherten signifikant mehr Kohlenstoff in der oberirdischen Biomasse als die
Kiefern(misch)bestände (Kategorien ÜK, ÜL). Die oberirdischen Biomasse-Kohlenstoffvorräte in den Wäldern der Kategorie N waren im Mittel
etwa doppelt so hoch wie in den jüngeren Kiefernreinbeständen, aber rund 35 % niedriger als der Durchschnitt der Buchen-Urwälder
(Kategorie OG; Tab. 3; Glatthorn et al. 2018;
Förster et al. 2021a). Einzelne Naturwälder der Kategorie N erreichten mit bis zu
228 t C ha − 1 aber sogar genauso hohe Werte wie die Buchen-Urwälder. In unseren Untersuchungsbeständen erwies sich die
Hauptbaumart in einem gemischten linearen Modell dabei als der wichtigste Faktor, aber auch im Laufe der Zeit nach einer Nutzungsaufgabe
erhöhten sich die Kohlenstoffvorräte (Förster et al. 2021a). Unsere Untersuchungen zeigten,
dass Kiefernreinbestände (Kategorie ÜL) jährlich etwa ein Viertel weniger oberirdische Biomasse produzierten als Laubwälder in natürlicher
Entwicklung.
Der Vergleich zwischen Laubwäldern (Kategorie N) und Kiefernbeständen (Kategorie ÜL) zeigte, dass auch die Biomasse und Produktivität
der Feinwurzeln in den Kiefernwäldern deutlich geringer waren als in den aus der Nutzung entlassenen Laubwäldern (Tab. 3; Förster et al. 2021b). Ein Einfluss der
Nutzungsaufgabe ließ sich bei diesen Kenngrößen allerdings nicht nachweisen. Der von uns untersuchte Kohlenstoffvorrat im Mineralboden war
unter den jüngeren Kiefernbeständen (Kategorie ÜL) größer als unter Buche (Kategorie N) (Diers et al.
2021).
|
|
Mittlere Kohlenstoffvorräte [t C ha−1]
|
|
Kompartiment
|
ÜL
|
ÜK
|
N
|
OG
|
Lebende Baumbiomasse oberirdisch | 70,0 | 83,0 | 142,0 | 192,0 |
Totholz | 1,6 | 3,3 | 4,6 | 35,0 |
Gehölzverjüngung | 0,4 | 0,3 | 1,0 | 1,5 |
Summe oberirdisch
|
72,0
|
87,0
|
148,0
|
228,0
|
Lebende Feinwurzeln (organische Auflage und 0 – 20 cm Mineralboden) | 0,2 | — | 1,2 | — |
Tote Feinwurzeln (organische Auflage und 0 – 20 cm Mineralboden) | 0,9 | — | 2,1 | — |
Kohlenstoffvorrat im Mineralboden | 141,0 | — | 99,0 | — |
Summe unterirdisch
|
142,0
| — |
102,0
| — |
Summe insgesamt
|
214,0
| — |
249,0
| — |
Nettoprimärproduktion oberirdisch pro Jahr | 3,3 | — | 4,3 | — |
Feinwurzelproduktion (0 – 20 cm Bodentiefe) pro Jahr | 0,4 | — | 0,7 | — |
N = natürliche Entwicklung, ÜK = Überführung kurzfristig, ÜL = Überführung langfristig,
OG = Old-Growth-Referenz |
Tab. 3: Mittlere Kohlenstoffvorräte sowie oberirdische Nettoprimär- und unterirdische Feinwurzelproduktion in den untersuchten
Waldkategorien N, ÜK und ÜL und in den Buchen-Urwäldern (Kategorie OG). Für weitere Informationen zu den Waldkategorien siehe
Tab. 1.
Quellen: Tabelle aus DBU 2020, verändert; Daten der Buchen-Urwälder aus Glatthorn et al. 2018, verändert.
Table 3: Average carbon stocks, aboveground net primary production and belowground fine root production in the studied forest
categories N, ÜK and ÜL and in the Slovakian primeval beech forests (category OG). For more information on the forest categories,
see Table 1.
Sources: table from DBU 2020, modified; data on primeval beech forests from
Glatthorn et al. 2018, modified).
3.3 Artenvielfalt
In den Wäldern mit natürlicher Entwicklung konnten wir die meisten (xylobionten) Pilz- und Käferarten erfassen (Tab. 2). Urwaldreliktarten der Käfer (Müller et al.
2005) wurden kaum nachgewiesen. Sowohl die Naturnähezeiger unter den Pilzen (Blaschke
et al. 2009) als auch die xylobionten Käfer zeigten eine Zunahme mit steigender Menge und damit auch mit steigender
Variabilität des Totholzes hinsichtlich Baumarten, Durchmesser und Zersetzungsgraden (Auswertungen dazu in Culmsee et al. 2021; Schneider, Karasch 2022).
Weiterhin konnten wir eine positive Reaktion der xylobionten Arten auf ein hohes Bestandsalter und eine lange Zeitdauer seit der
Nutzungsaufgabe nachweisen. Im Gegensatz zu Pilzen und Käfern wiesen Gefäßpflanzen in unseren Untersuchungen die geringsten Artenzahlen in
den Wäldern der Kategorie N auf. Waldtypische Gefäßpflanzen (Schmidt et al. 2011) und
epiphytische Moose und Flechten waren dort dagegen häufiger vertreten.
4 Diskussion
Auf Grundlage der von uns erhobenen Daten sowie der Referenzdaten aus Buchen-Urwäldern konnte der OGI als Maß für die Ähnlichkeit der
Waldstruktur mit reifen Waldentwicklungsphasen abgeleitet werden. Dies ermöglicht die Beurteilung der Ähnlichkeit der Waldstruktur für
jeden einzelnen Untersuchungsbestand. Für Waldbestände der Kategorie N wurden im Durchschnitt höhere OGI-Werte berechnet als für die
Kiefern(misch)wälder der Kategorien ÜK und ÜL. Dennoch fehlen in den N-Beständen die für Old-Growth-Bestände typischen Merkmale,
insbesondere eine große Totholzmenge und -diversität, höhere Baumdimensionen, eine größere Anzahl verschiedener Waldentwicklungsphasen und
eine ausgeprägte Naturverjüngung. Es zeigte sich, dass sich die Untersuchungsbestände im Zuge der natürlicher Waldentwicklung den
Old-Growth-Beständen hinsichtlich ihrer Waldstruktur annähern. Ohne größere Störungsereignisse wird sich der Eintritt des Hauptbestands in
die Zerfallsphase sowie die nachfolgende Bestandsverjüngung jedoch noch Jahre bis Jahrhunderte hinziehen (Jönsson et al. 2009; Svoboda et al. 2010). Momentan
befinden sich unsere Untersuchungsbestände größtenteils in der Optimalphase ohne Störungen und mit einem geringen Lichtangebot. Das
Totholzangebot und damit einhergehend auch die Variabilität des Totholzes (Baumarten, Durchmesserstärken, Zersetzungsgrade) fällt in
diesen Wäldern im Vergleich zu Buchen-Urwäldern im Old-Growth-Stadium eher gering aus (Kunttu et al.
2015).
Der Vergleich der Kohlenstoffvorräte in der oberirdischen Biomasse und der Nettoprimärproduktion in den heimischen
Untersuchungsbeständen mit den Buchen-Urwäldern weist auch in Bezug auf die funktionalen Kenngrößen der Kohlenstoffspeicherung und
-sequestrierung auf eine Annäherung der aus der Nutzung entlassenen Laubwälder an die Reifephase von Buchen-Urwäldern hin. Zwar hatte die
Hauptbaumart den größten Einfluss auf die Kohlenstoffvorräte in der oberirdischen Biomasse, diese nahmen aber auch nach der
Nutzungsaufgabe im Laufe der Zeit zu und zeigten, dass nach Nutzungsaufgabe Biomasse und damit Kohlenstoffvorräte bis zum Beginn
natürlicher Störungen weiter ansteigen können.
Aufgrund fehlender Referenzdaten aus Urwäldern ist eine zur Waldstruktur analoge Beurteilung von Waldbodenvegetation, Pilz- und
Käferarten bislang nicht möglich. Auch spielen abweichende Umweltvariablen (Höhe, Temperatur, Niederschlag, Bodenbedingungen) in den
Referenzbeständen bei der Beurteilung der Artenvielfalt eine größere Rolle. Eine Einordnung im Vergleich zu den Buchen-Urwäldern konnte
daher nicht vorgenommen werden. Grundsätzlich gilt, dass ein reiches und vielfältiges Totholzangebot die Voraussetzung für eine hohe
Diversität an xylobionten Pilzen und Insekten darstellt (Heilmann-Clausen, Christensen 2004;
Brin et al. 2011) – dies bestätigen auch die Ergebnisse aus den von uns
untersuchten Waldbeständen (Culmsee et al. 2021). Die im Gegensatz dazu stehende
geringere Artenvielfalt in der Bodenvegetation in Wäldern mit natürlicher Entwicklung zeigen auch Vergleichsuntersuchungen von
Naturwaldreservaten und Wirtschaftswäldern (Meyer 2018). Durch fehlende Störungen in den
Wäldern der Optimalphase ist das Lichtangebot flächendeckend eher gering. Der Wert nutzungsfreier Wälder für die Bodenvegetation gründet
sich vielmehr auf einer höheren Anzahl schattentoleranter, waldtypischer Gefäßpflanzen (Schmidt et al.
2011).
5 Fazit
In einem so stark durch den Menschen beeinflussten Naturraum wie Mitteleuropa ist es schwierig, die Struktur, Artenzusammensetzung
und Funktion ursprünglicher Waldlebensräume zu definieren. Essenziell bei der Bewertung der Naturnähe von Wäldern sind daher die zugrunde
gelegten Referenzbestände. Der im WiNat-Projekt entwickelte OGI erlaubt eine Naturnähebewertung der Struktur reifer Wälder auf Basis
ähnlicher Daten aus Buchen-Urwäldern. Auch die Kohlenstoffspeicherung lässt sich durch einen Vergleich mit Werten aus Buchen-Urwäldern in
Bezug auf die erreichbaren Kohlenstoffvorräte in reifen Buchenwäldern einordnen. Die weiterhin erfassten Biodiversitätskenngrößen sind
jedoch auch ohne Referenzierung von großer Bedeutung und geben weiterführende Hinweise auf die Naturnähe von Wäldern, z. B. über das
Vorkommen von Arten mit enger Waldbindung oder von bereits in der Literatur verifizierten Indikatorarten.
Der relative Einfluss der Nutzungsaufgabe, der Baumartenzusammensetzung und des Bestandsalters auf die untersuchten Kenngrößen ließ
sich nur vereinzelt identifizieren. Es zeigte sich jedoch, dass der OGI mit zunehmender nutzungsfreier Zeit ansteigt. Eine
naturnahe Ausstattung der Waldstruktur ist wiederum Voraussetzung für das Auftreten weiterer Naturnähezeiger wie
spezialisierter Pflanzen-, Flechten-, Pilz- oder Insektenarten. Nicht untersucht wurde, in welchem Umfang Wälder jüngerer Sukzessions- und
Entwicklungsstadien in unserer Landschaft natürlicherweise eine Rolle spielen würden. Es bestehen noch große Wissenslücken über Bedeutung
und Auswirkungen größerer natürlicher Störungen wie Sturm oder Feuer in temperaten Wäldern.
6 Literatur
↑
Bauhus J., Puettmann K., Messier C. (2009): Silviculture for old-growth attributes. Forest Ecology and Management
258(4): 525 – 537.
↑
Blaschke M., Helfer W. et al. (2009): Naturnähezeiger – Holz bewohnende Pilze als Indikator für Strukturqualität im Wald. Natur und Landschaft 84(12):
560 – 566.
↑
BMU/Bundesministerium für Umwelt, Naturschutz und Reaktorsicherheit (Hrsg.) (2007): Nationale Strategie zur biologischen
Vielfalt. BMU. Berlin: 178 S.
↑
Brin A., Bouget C. et al. (2011): Diameter of downed woody debris does matter for saproxylic beetle assemblages in
temperate oak and pine forests. Journal of Insect Conservation 15(5): 653 – 669.
↑
Culmsee H., Schneider H. et al. (2021): WiNat – Wildnis Naturerbe. Naturwald-Entwicklung und Wildnisgebiet-Umsetzung im Nationalen
Naturerbe. Bundesprogramm Biologische Vielfalt. Förderschwerpunkt Ökosystemleistungen. DBU Naturerbe GmbH. Osnabrück:
96 S.
↑
DBU/Deutsche Bundesstiftung Umwelt (Hrsg.) (2020): Wildnis Naturerbe – Wie können wir Naturnähe im Wald messen? Ergebnisse des Projekts „Wildnis Naturerbe“
(WiNat) – Naturwald-Entwicklung und Wildnisgebiet-Umsetzung im Nationalen Naturerbe. DBU. Osnabrück: 38 S.
↑
DBU Naturerbe GmbH (2014):
Naturnahe Waldentwicklung auf DBU-Naturerbeflächen. Grundsätze zur Entwicklungssteuerung. Stand: 1.12.2014. DBU Naturerbe GmbH.
Osnabrück: 8 S.
↑
Diers M., Weigel R. et al. (2021): Soil carbon and nutrient stocks under Scots pine plantations in comparison to European beech
forests: A paired-plot study across forests with different management history and precipitation regimes. Forest Ecosystems 8:
47.
↑
Engel F., Meyer P. et al. (2019): Wälder mit natürlicher Entwicklung in Deutschland. AFZ DerWald 13: 30 – 33.
↑
FAO/Food and Agriculture Organization (2020): Global forest resources assessment 2020: Main report. FAO. Rome: 184 S. DOI:
10.4060/ca9825en
↑
Feldmann E. (2018): Forest structure and structural dynamics of virgin beech forests in Slovakia.
Dissertation. Georg-August-Universität Göttingen. Göttingen: 132 S.
↑
Feldmann E., Glatthorn J. et al. (2018): A novel empirical approach for determining the extension of forest
development stages in temperate old-growth forests. European Journal of Forest Research 137(3): 321 – 335.
↑
Förster A., Culmsee H., Leuschner C. (2021a): Thinned northern German Scots pine forests have a low carbon storage
and uptake potential in comparison to naturally developing beech forests. Forest Ecology and Management 479: 118575.
↑
Förster A., Hertel D. et al. (2021b): Belowground consequences of converting broadleaf to conifer forest: Comparing the fine root systems of
European beech and Scots pine. Forest Ecology and Management 496: 119457.
↑
Glatthorn J. (2018): Structure, productivity and carbon storage of primeval European beech forests.
Dissertation. Georg-August-Universität Göttingen. Göttingen: 128 S.
↑
Glatthorn J., Feldmann E. et al. (2018): Biomass stock and productivity of primeval and production beech forests: Greater canopy structural
diversity promotes productivity. Ecosystems 21(4): 704 – 722.
↑
Goff F.G., West D. (1975): Canopy-understory interaction effects on forest population structure.
Forest Science 21(2): 98 – 108.
↑
Heilmann-Clausen J., Christensen M. (2004): Does size matter? On the importance of various dead wood fractions for
fungal diversity in Danish beech forests. Forest Ecology and Management 201(1): 105 – 117.
↑
Höltermann A., Jessel B. et al. (2019): Wälder im Klimawandel: Steigerung von Anpassungsfähigkeit und Resilienz durch mehr Vielfalt und
Heterogenität. Ein Positionspapier des Bundesamtes für Naturschutz. BfN. Bonn: 31 S.
↑
Jönsson M.T., Fraver S., Jonsson B.G. (2009): Forest history and the development of old-growth characteristics in fragmented boreal
forests. Journal of Vegetation Science 20(1): 91 – 106.
↑
Kahl T., Bauhus J. (2014): An index of forest management intensity based on assessment of harvested tree volume, tree species composition and
dead wood origin. Nature Conservation 7: 15 – 27.
↑
Korpel' Š. (1995): Die Urwälder der Westkarpaten. Fischer. Stuttgart: 310 S.
↑
Kowarik I. (1999): Natürlichkeit, Naturnähe und Hemerobie als Bewertungskriterien. In: Konold W., Böcker R.,
Hampicke U. (Hrsg.): Handbuch Naturschutz und Landschaftspflege. Kap. V-2.1. Ecomed. Landsberg: 18 S.
↑
Kunttu P., Junninen K., Kouki J. (2015): Dead wood as an indicator of forest naturalness: A comparison of methods.
Forest Ecology and Management 353: 30 – 40.
↑
Leuschner C. (2015): Monospecific and mixed stands of Fagus and Nothofagus species in
the temperate zones of the world. Berichte der Reinhold-Tüxen-Gesellschaft 27: 49 – 63.
↑
Majdi H. (1996): Root
sampling methods – Applications and limitations of the minirhizotron technique. Plant Soil 185: 255 – 258.
↑
Markgraf F. (1931): Aus den südosteuropäischen Urwäldern. Die Wälder Albaniens. Zeitschrift für das
Forst- und Jagdwesen 73(1): 1 – 32.
↑
Meyer P. (2018): Wald ohne
Bewirtschaftung: Sinnvoll oder überflüssig? AFZ DerWald 20: 26 – 29.
↑
Meyer P., Aljes M. et al. (2021): Quantifying old-growthness of lowland European beech forests by a multivariate indicator for forest
structure. Ecological Indicators 125: 107575.
↑
Meyer P., Brößling S. et al. (2013): Monitoring von Waldstruktur und Vegetation in hessischen
Naturwaldreservaten. Stand: April 2013. Nordwestdeutsche Forstliche Versuchsanstalt. Göttingen: 63 S.
↑
Müller J., Bußler H. et al. (2005): Urwald relict species – Saproxylic beetles indicating structural qualities and habitat tradition.
Waldökologie Online 2: 106 – 113.
↑
Müller-Using S., Bartsch N. (2009): Decay dynamic of coarse and fine woody debris of a beech (Fagus sylvatica
L.) forest in Central Germany. European Journal of Forest Research 128: 287 – 296.
↑
NFP/Niedersächsisches Forstplanungsamt (2001): Anweisung zur Betriebsinventur. Unveröff. NFP.
Wolfenbüttel: 88 S.
↑
Persson H. (1980): Fine-root dynamics in a Scots pine stand with and without near optimum nutrient and
water regimes. Acta Phytogeographica Suecica 68: 101 – 110.
↑
Peters R. (1992): Ecology of beech forests in the northern hemisphere. Dissertation. Universität
Wageningen. Wageningen: 122 S.
↑
Sabatini F.M., Burrascano S. et al. (2018): Where are Europe's last primary forests? Diversity Distribution 24(10):
1.426 – 1.439.
↑
Schall P., Ammer C. (2013): How to quantify forest management intensity in Central European forests. European
Journal of Forest Research 132(2): 379 – 396.
↑
Schmidt M., Kriebitzsch W.-U., Ewald J. (2011): Waldartenlisten der Farn- und Blütenpflanzen, Moose und Flechten Deutschlands.
BfN-Skripten 299: 111 S.
↑
Schneider H., Karasch P. (2022): Diversität holzbewohnender Pilze in nutzungsfreien Buchenwäldern. AFZ DerWald 3:
30 – 34.
↑
Svoboda M., Fraver S. et al. (2010): Natural development and regeneration of a Central European montane spruce
forest. Forest Ecology and Management 260(5): 707 – 714.
↑
Swanson M.E., Franklin J.F. et al. (2011): The forgotten stage of forest succession: Early-successional ecosystems on forest sites.
Frontiers in Ecology and the Environment 9(2): 117 – 125.
↑
Westphal C., Tremer N. et al. (2006): Is the reverse J-shaped diameter distribution universally applicable in
European virgin beech forests? Forest Ecology and Management 223(1 – 3): 75 – 83.
↑
Winter S., Fischer H.S., Fischer A. (2010): Relative quantitative reference approach for naturalness assessments of forests. Forest Ecology and
Management 259: 1.624 – 1.632.
↑
Wirth C., Gleixner G., Heimann M. (2009): Old-growth forests – Function, fate and value. Ecological
Studies 207. Springer. Berlin: 540 S.
Förderung
Das Projekt „WiNat – Wildnis Naturerbe“ wurde im Rahmen des Bundesprogramms Biologische Vielfalt in der Fördermaßnahme „Forschung zur
Umsetzung der Nationalen Strategie zur biologischen Vielfalt (F & U NBS)“ von 2014 bis 2020 durch das Bundesministerium für Bildung
und Forschung (BMBF) und das Bundesamt für Naturschutz (BfN) mit Mitteln des Bundesministeriums für Umwelt, Naturschutz und nukleare
Sicherheit (BMU) gefördert.
Zusatzmaterial zum Beitrag
Schneider H. et al.: Wie kann Naturnähe von Wäldern bewertet werden. Natur und Landschaft 98(2): 49 – 57. DOI:
10.19217/NuL2023-02-01
Zusatzmaterial (Bootstrapping zur
Variablenselektion des Old-Growth-Indikators (OGI), Abb. A, Tab. A, B, C, Literatur) als PDF herunterladen (PDF 945 kB)

